Mac Vet Rev 2014; 37 (1): 5 - 34
 10.14432/j.macvetrev.2014.09.022
10.14432/j.macvetrev.2014.09.022
Received: 07 September 2013
Received in revised form: 12 July 2019
Accepted: 30 September 2013
Available Online First: 15 September 2013
Published on: 15 March 2014
Keywords: boar; semen storage; artificial insemination; fertility
AI is of great economic value for the pig industry. In the past two decades, there has been an enormous increase in the development of field AI services in the majority of countries concerned with pork production. In the authors’ opinion, several deciding factors have paved the way for many producers to use AI in the breeding management programs (1). First, the industry’s requirement for improved leanness and maternal traits has stimulated the need for changes in genetic programs, and AI provides a viable tool for this change. Second, increased understanding of the merits of utilizing sires with predictable superior genetics. Third, favorable field results have been reported after AI with liquid-stored semen. Fourth, on-farm AI as compared with natural service can provide substantial economic benefits, especially in larger production units. Fifth, semen is available from a number of commercial sources at a competitive price. Sixth, AI technique is simple to learn and the equipment cost is reasonable.
More than 99% of inseminations conducted worldwide are made with liquid-stored semen at 15-20°C for 0-5 days, with 85% of all inseminations are carried out on the day of collection or on the following day for meat production market. Less than 1% of all inseminations are made with cryopreserved semen for upgrading the genetic nucleus in a particular country or herd. The commercial use of liquid-stored semen includes do-it-yourself AI (DIY AI; pig producers collect semen and inseminate the females on their own farm, i.e. on-farm semen collection, processing and AI) and extensive networks of AI centers with services set up to deliver the semen (2). The numbers of AI centers in Germany, Spain, Denmark, Poland, Belgium, Sweden and Norway are 22, 74, 13, 22, 37, 3 and 1, respectively (~172 centers in Europe). AI with liquid-stored semen has reached a very high level in USA and Mexico at ~ 90% and in Canada at 80%. In South American countries, pig production has greatly increased as a total in the last decade (Brazil, Argentina and Chile by 450, 112 and 65%, respectively). This features an extremely high AI level with almost 100% in Chile and 66% in Brazil. China represents an example for the increases in pig production, with ~ 33 million tons of pork have been produced in China in 1996. In 2008, production was more than 47 million tons. During the next years, further growth of pork production is predicted in China and Brazil (3).
The aim of the present review is to shed some light on the porcine AI industry. By the end of this review, the reader should be able to understand: (a) the current status of swine AI in the world, (b) significance and limitation of AI with liquid and frozen semen, (c) the biological traits of porcine semen in relation to in vitro sperm storage, (d) the criteria used for selection of a boar stud as a semen supplier, (e) how to process boar semen for liquid and frozen storage in the commercial settings, and (f) how to improve fertility and prolificacy of boar semen after AI.
1. North American AI centers
Boar studs with 100-500 sires are most common in Canada and USA. Most boars are housed in stalls and fed according to their body condition or weight. The predominant age of boars is 1-2 years. Evaporative and mechanical systems are the predominant methods of thermal control. The annual boar replacement (culling) rates ranges between 20 and 70%. The primary reasons for culling include genetic improvement, semen quality and feet and leg problems. Training of boars for semen collection is initiated at 8-9 months of age and requires 1-3 weeks to complete. Collection occurs commonly on Monday and Thursday and boars are rested 3-7 days between collections. Most studs utilize double gloves with the outer pair discarded prior to semen collection. The most common type of collection containers used is a disposable bag inside a thermal container. Pre-warming collection containers is practiced by 75% of studs. The average sperm produced/boar/week is 51-150 billions resulting in 21-40 doses/boar/week. Ejaculate pooling is performed in 60% of studs, using 2-3 or 4-6 ejaculates. Evaluation of semen for sperm motility is done within 0-5 min of warming in an extender with viewing at 100-400 x magnification. Sperm concentration is determined by photometers and CASA for 88% of studs. Ejaculates are discarded most frequently at a rate of 1-10% due to poor motility, abnormal sperm morphology, bacteriospermia and other factors involving low sperm concentration, low semen volume, presence of contaminants such as blood, urine and off odor or abnormal color of semen. The majority of discards occur as a result of individual boar and season. Semen culture for bacterial growth in ejaculates is performed routinely on set schedules in 84% of studs, and only in cases of problems for 14% and is not performed in 2% of studs. For quality control, most studs maintain semen samples following extension for 3-4 or 3-7 days (4, 5).
The majority of studs use proprietary commercial extenders followed by non-proprietary BTS (Beltsville Thawing Solution) extender. The relative contribution of each storage medium to all studs is as follows: 50% Androhep®, 30% X-Cell®, 25% BTS, 20% Enduraguard®, 12% Geddil™, 10% Preserv™, 7% Ivo-Zeist™ and MR-A®, and 3% Acromax®, Cell-Lution™, Safe-Cell™, Survivor™ and Vital®. The choice of an extender is based primarily on the duration of preservation for sperm fertility, the cost and the brand recognition in 65%, 51% and 33% of studs, respectively. When preparing the extender, 74% of the studs use an in-house water purification system while 26% of them purchase purified water. Water quality is tested for impurities by 67%, for microbes by 58% or it is reported to occur by assurance of the water manufacturer by 33% or is reported as unknown by 7% of studs. Semen is packaged at 2-4 billion fertile sperm cells/dose in > 90% of studs. Of studs, 42% indicate that they alter the number of sperm in the final packaged dose based on breed or genetic line. For the final sperm numbers in a dose, most studs indicate that they adjust the total cells in the dose based on morphologically normal or viable sperm. For the final packaging, sealed tubes or bags containing 60-80 ml of diluted semen are used in most studs. Custom processing for packaging or for specific volumes is not a service provided by 81% of studs, but in those that do, some process for packaging in the GEDIS® device (IMV technologies, France) and others process for frozen semen. Semen delivery is accomplished by stud personnel for 45% of studs while 36% of studs use an independent courier and 27% use another system which includes a company courier that has delivery duties but are not stud related (5).
2. Dutch AI centers
In Europe, the pig AI rate is between 25 and 98%. In The Netherlands, the pig AI rate is 98%, based on the annual 4.3 million sow matings. About 97% of the insemination doses are purchased from AI companies of which 90% through the cooperative pig AI centers Varkens KI Nederland (contain ~ 1800 boars producing 3.7 million inseminations per year) and Varkens Kl Twenthe. Only 3% ‘on-farm AI’ is used. Boars located on the AI centers originate from several breeding companies such as TOPIGS (94.9%), PIC (3.4%), Nord West (0.6%), Danbred (0.3%), JSR (0.1%) and others (0.7%). In contrast to a lot of other countries, where inseminations are alternated with natural mating within a cycle of a sow, AI is solely used in The Netherlands. The Dutch were the first to introduce the pooling of semen (since 1992) for commercial herds, however since the major outbreak of Classical Swine Fever in 1997, where two pig AI stations were involved as well, the use of pooled semen was prohibited by the Dutch government. Semen production for commercial use in The Netherlands is only allowed on EU approved semen collection centers (Directive 90/429/EEC). All centers of Varkens KI Nederland are under a strict hygienic program, in which critical points in the production are tested on a daily, weekly and monthly schedule for bacteriological contamination. The working methods at the AI stations are standardized and controlled during the whole production procedures (6, 7).
Boar ejaculates are collected in separate collection pens on dummy sows using the “gloved-hand” technique (sperm-rich fraction) or in an automated semen collection system (Collectis®, IMV technologies). Before semen collection, body temperature is measured to ensure the health status of the boar on the collection day. The semen is transported from the collection area to the lab via a pneumatic tube system. At the AI lab, the semen is extended with a two-step dilution method. The volume of the ejaculate (ml) is diluted with ~ the same volume (ml) of Solusem® short-term (≤ 72 h) extender at 32°C within 15 min after collection (step 1). After this dilution, semen quality is assessed by measuring sperm motility (fresh, 24 and 72 h after production) and density using CASA system. Each fourth to sixth ejaculate of a boar is subjectively assessed for sperm longevity and morphological abnormalities (fixed-stained samples) using a phase-contrast microscope. However, when aberrations are observed or when otherwise the ejaculate quality is doubted, each ejaculate of that boar will be evaluated for sperm longevity and morphology. Because morphological evaluations are time-consuming, the results are only available after distribution of semen doses. After semen quality assessment, semen is finally diluted (step 2) with Solusem® 20°C to a minimum of 1.5 billion sperm in 80 ml, packaged in tubes via a full automatic filling device and then stored in a temperature-conditioned room at 17°C (6).
Extender production is performed on-lab. Therefore, modern water production equipment is available. Water demineralization is performed by reverse osmosis and disinfection is done by UV-lights. The quality of both production water and prepared extender is checked by measuring pH, conductivity, osmolarity and bacteriology. For the transport of insemination doses to the client, temperature-conditioned cars are used. The transport temperature is 17°C. In The Netherlands almost all pig breeders have temperature controlled boxes or cabinets to store the doses until use on the farm (6).
Varkens KI Nederland records boar- and ejaculate-related data such as boar identification (tattoo # and name), genetic line and age of the boar (25±12 months), days between ejaculation (4.5±2.5 days), AI station, location and date of ejaculate production, semen collector, laboratory where ejaculate is processed, laboratory technician who assessed semen quality, volume and concentration of the ejaculate (84±11x109 sperm per ejaculate), motility (fresh, 24 and 72 h), morphological abnormalities, concentration of the insemination dose and number of doses produced per ejaculate (35±15x106) in the AI database. At the farms in The Netherlands, the number of inseminations per cycle is 1.6 with a range of 1-3 and ~ 10-15 sows are inseminated from semen of one ejaculate (7).
3. Belgian AI centers
The size of the herds in the centers varies between 4 and 165 boars, with an average of 45 boars. Semen doses contain at least 3 billion spermatozoa in 100 ml extended semen. Sperm morphology and motility are considered the most important parameters for sperm quality evaluation. Lower limits of 80% normal spermatozoa and 70% motility are used. Motility assessment is performed visually in all but one of the AI centers. Ninety-seven percent of AI centers use a short-term semen extender (storage for 3 days). Since semen transport times are short in Belgium, there is no necessity for long-term storage of diluted porcine semen. In 50% of the AI centers, long-term extenders (storage for 7 days) are used on a part of the collected semen, mainly for storage over the weekend. In some centers, extenders are added to the ejaculate at room temperature, without negative effects on fertility. Belgian porcine AI centers generally incorporate semen quality examinations in their daily routine, especially for motility assessment. In minority of AI centers, especially the smaller ones, semen morphology examination is performed more systematically. This focus on semen quality, together with the consistent use of a sufficient number of spermatozoa per semen dose, reflects the concern of Belgian centers to produce top quality semen doses (8, 9).
1. Liquid semen
Current procedures of pig AI employ the extension of fresh whole ejaculate gel-free semen with media which guarantee a fertile shelf life between 3-5 days and the use of 2.5-4 billion spermatozoa per insemination in a large volume of liquid-stored semen (70-100 ml) deposited intracervically two or three times during the estrous period. These conditions limit the number of doses that can be prepared from one ejaculate to ~ 20-25. To increase the efficient use of spermatozoa of boars of high genetic merits, several strategies have been proposed, including reducing the number of inseminations per estrous period to one insemination only (by increasing the fertile lifespan of sperm cells or improving the prediction of ovulation time) and utilizing new AI techniques, which allow a change in the site of semen deposition such as post-cervical or deep intrauterine insemination (10, 11).
At the practical level, semen storage media can be divided into two major groups: those designed for short-term preservation (1-3 days; examples: BTS & Vital®) and media for long-term preservation (> 3 days; examples: Androhep®, X-Cell®, MULBERRY III®, MR-A® and Acromax®). The former are mainly used in short distance semen dose distribution networks (such as European systems in which semen doses are frequently produced at the farm itself) while long-term media are generally used in programs such as those in USA and Norway, where the site of semen production is far away from the site of insemination. The advantages of long-term media are: long distance transport, conducting diagnostic tests (PCR and semen evaluation) and improving the organization of tasks at semen collection centers. For a planned semen storage time < 72 h, it is preferable to use a short-term medium as it is less expensive and has been associated with a similar reproductive outcome to that of the long-term one. When semen doses are needed to be stored > 72 h, it is recommended to use a long-term medium on a higher sperm concentration to compensate for reduced sperm viability due to in vitro aging of spermatozoa (12).
One of the most common media is BTS, but many companies commercialize different kinds of media as mentioned above (13). The main limitation of these media is the preservation of semen at 15-18°C in special temperature-controlled boxes. This temperature represents a logistic problem not only for the international transportation but also for the on-farm storage. Doubtless, the possibility of preserving semen at a temperature below 15°C would help reduce sperm metabolic activity and protect the semen from detrimental effects of microbial contamination. Bacteriospermia is a frequent finding in porcine semen stored in liquid media for 3-7 days before AI (14). Nevertheless, storage of boar semen at 5-8°C for > 24 h decreases the conception and embryo survival rates (15, 16). Semen stored at 15-18°C has been found more successful based upon farrowing rates and litter size (12, 17).
2. Frozen semen
Continued interest in swine frozen semen is based upon its success in the dairy industry. While one bull can produce enough AI doses to serve ≥ 40,000 cows per year, one boar produces < 40 doses per week or enough doses to serve < 2000 sows per year. Furthermore, most cattle receive a single AI with 15 million sperm per dose, while most pigs receive a double insemination of 2.5-3 billion sperm in each of two intracervical AI doses. In swine, the need for a greater number of spermatozoa may be important for achieving high fertilization rates in response to multiple ovulations, the dilute ejaculate, long uterine horns and extended interval from AI to ovulation (18). Frozen semen has an unlimited durability than liquid-stored semen and can be used in breeding sows for the genetic improvement at remote regions. Nonetheless, cryopreservation of boar semen is still expensive, complex and laborious production process. In comparison with liquid semen, the low fertility and prolificacy (2-3 piglets less) after AI with frozen-thawed semen limit its use in the international markets (19).
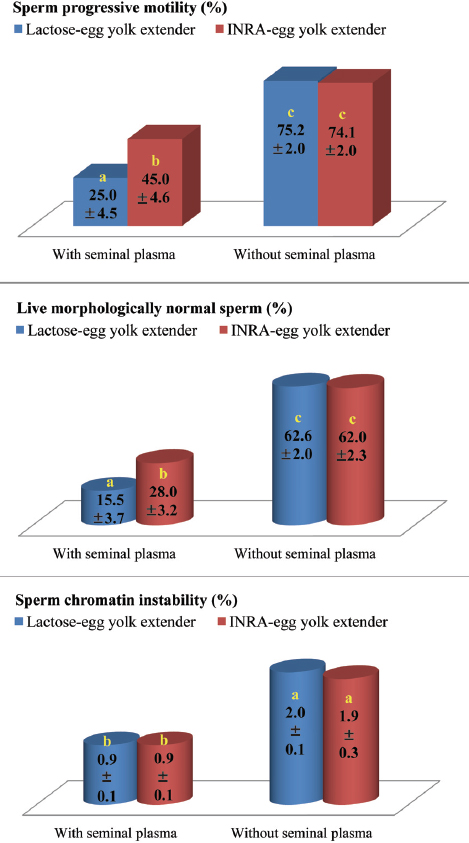
Frozen semen is presently used for the international exchange among nucleus farms to help maintain genetic diversity and to reduce the risk of disease transmission associated with boar and liquid semen entry. The use of frozen semen beyond several days can provide increased flexibility for on-farm use and allow additional time for disease tests. There may be a potential use of frozen semen for short-term banking during periods of low demand and while sires await breeding value tests. Opportunities exist for use of frozen semen in long-term banking of sire lines, creation of semen pools for genetic progress evaluation and for use in emergencies (20). The current procedure for AI with frozen semen involves the deposition of a high number of thawed spermatozoa (5-6 billion) extended in 80-100 ml of diluent into the cervix. However, recent findings in our lab show that post-thaw dilution of frozen boar semen causes deterioration of sperm quality characteristics (Tables 1-3) (21). In fact, the development of a low-volume deep intrauterine insemination technique has decreased not only the number of frozen sperm needed for successful AI to ≤ 1 billion but also the detrimental post-thawing ‘dilution effect’ on spermatozoa (21). This technique coupled with insemination of sows in spontaneous estrus close to the time of ovulation (0-4 h before ovulation; checked by ultrasonography) have improved, to a great extent, the farrowing rate (81.4%) and litter size (9.6 piglets) of frozen semen (10, 19, 22).
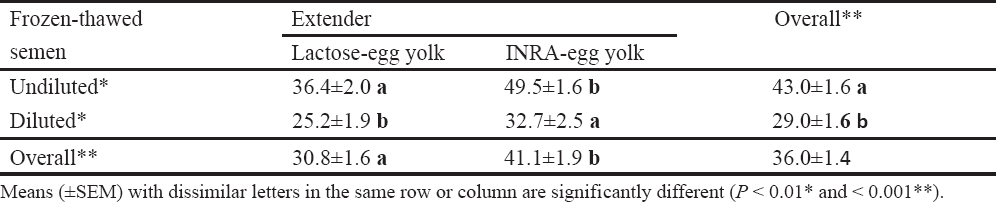
Table 1. Influence of extender and post-thawing dilution of frozen boar semen on proportions of sperm progressive motility (21).
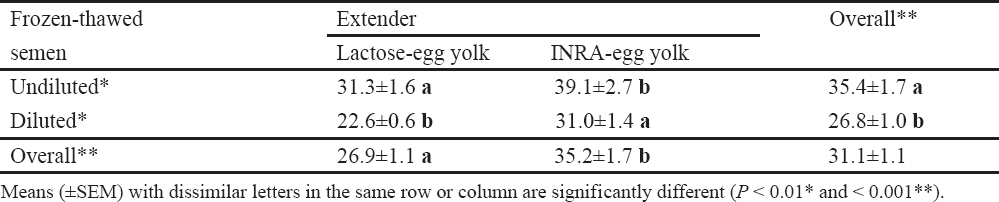
Table 2. Influence of extender and post-thawing dilution of frozen boar semen on proportions of live morphologically normal spermatozoa (21).
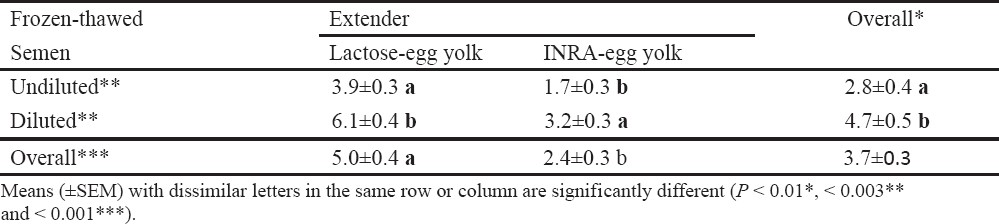
Table 3. Influence of extender and post-thawing dilution of frozen boar semen on incidence (%) of sperm chromatin instability (21).
Spermatozoa, whether present in whole semen or washed suspensions, are comparatively ineffective ‘anaerobes’. Unlike those of the human, bull, ram and billy goat, spermatozoa of the boar cannot obtain sufficient energy for powering their motility (total and progressive) from the anaerobic breakdown of fructose or glucose alone. On the contrary, boar spermatozoa are highly effective ‘aerobes’ and their respiratory coefficient equals that of the bull and ram spermatozoa. The presence of oxygen and sufficient oxidizable energy substrates is essential for maintenance of sperm motility in vitro (23, 24). Aerobic incubation of buffered whole semen at 37°C for 3 h causes a complete loss of sperm motility, indicating that boar semen contains insufficient energy substrates and addition of oxidizable substrates is necessary to maintain sperm motility in vitro (23). Boar spermatozoa can metabolize aerobically fructose, glucose, mannose, glycerol, lactic acid, pyruvate, citrate and acetic acid. They are unable to oxidize sorbitol and inositol, presumably owing to absence of the necessary enzyme system. Thus, they differ from ram spermatozoa which readily metabolize sorbitol to fructose using a highly active sorbitol dehydrogenase (23-29).
The high ratio of unsaturated: saturated fatty acids in the phospholipids and the low cholesterol content of plasma membrane may contribute to the extreme sensitivity of boar sperm to cold shock (2, 30). Therefore, delivering cholesterol to the plasma membrane of boar spermatozoa is associated with increasing their resistance against cold stress (18, 31, 32), although in a recent study, this treatment does not improve the fertility of frozen semen (33). Spermatozoa of whole ejaculate are more susceptible to cold shock than those of sperm-rich fraction, due to presence of a cold shock sensitizing factor in secretions of boar accessory sex glands (34, 35). Incubation of diluted semen above 15°C increases sperm resistance to cold shock (36-38). The extreme vulnerability of boar spermatozoa to cold shock means that in practice, semen samples should be kept at 15-20°C, since a further decrease in the storage temperature below 12°C compromises their viability (39, 40). Nonetheless, the need to preserve boar semen at these moderately reduced temperatures restricts their storage capacity since cell metabolism cannot be slowed down (34, 41) and microbial conditions may not be as effectively controlled as at lower temperatures (5-10°C) (42).
The basic requirements for a swine AI program are boar (or semen) selection and purchase, training of boars for semen collection, semen evaluation, processing and storage and estrus detection and insemination. Semen is collected by the “gloved-hand” technique from boars at a frequency of two times per week (43-45). The standard procedure involves collection of the whole ejaculate, a modified whole ejaculate or the sperm-rich fraction (34). Currently, the use of an automated semen collection system (Collectis®, IMV Technologies) has not only maximized the labor and collection efficiency but also reduced the incidence of bacterial contamination in extended semen. The Collectis® system regulates air pressure and vacuum (similar to those found in the milking palor) to control the volume and pressure of air supplied to an artificial vagina which holds the boar’s penis during collection (46).
In view of our experience, the main criteria for selection of a boar stud as a reliable semen supplier are: (a) having a documented quality control program in place, (b) providing a high level of biosecurity in the animal area and the AI laboratory, (c) identifying and discarding low quality ejaculates, (d) having well-trained personnel for collection and processing of semen, (e) using high quality water and high quality semen preservation media, (f) using a medium which is appropriate for the duration of semen storage, (g) providing a label on each dose of semen that includes relevant information, such as collection date, sire, identity of semen pool and expiration date, (h) keeping a reference sample of each ejaculate or pool, (i) documenting results of semen evaluation on reference samples, and (j) providing a proper temperature control during shipment of semen doses.
A semen extender is the backbone of the insemination dose. It provides the semen with nutrients and keeps the pH at the optimum level. The composition of the extender is important for semen longevity (2). Depending on the country, a longer shelf life is important in order to enable long distance transportations (3). In general, costs related to reproduction represent a low proportion of the overall cost of pig production and have been estimated at 1.9%. If we consider that most of this cost is associated with the human resources needed for estrus detection, semen collection and AI, the storage extender represents an insignificant portion of the complete production costs compared with its serious economic consequences on farrowing rates and litter sizes (12). Owing to high personnel costs, most of the labs use powdered preparations or concentrated liquid stock solutions that only require dilution in distilled water (5). This method contrasts with the in-house preparation, which involves separately weighing each component before dilution (6). In most cases, the testicular tissue and accessory glands of the boar are bacteria-free and therefore bacterial contamination of the ejaculate occurs during the semen collection process. An antibiotic should be added to the extender since its components (glucose) and the storage temperature (15-18°C) promote the growth of gram negative bacteria such as E.coli, Salmonella and Pseudomonas species. Bacterial contamination of stored semen causes diminished sperm motility, sperm agglutination or clumping, damaged sperm acrosomes and reduction of pH to the acidic levels (5.7-6.4). Thus, adding antibiotics at the appropriate concentration coupled with improving the hygienic measures during semen collection and processing can ameliorate preservability and fertility of semen (14, 42, 47-52). Penicillin plus streptomycin (1 g/L), which still have some importance in suppressing leptospira, were initially the combination most frequently used. Thereafter, aminoglycosides are still successfully used including gentamicin sulfate (0.2-0.3 g/L), neomycin sulfate (1 g/L), kanamycin sulfate (0.1-0.2 g/L) and dibekacin (0.1 g/L). Most recently, new generation of antibiotics (ceftiofur and apramycin) are being used, though no conclusive results are available yet (2, 12, 53, 54). Furthermore, since gram negative bacteria are the predominant type detected in boar semen, the endotoxins released after inhibition of these bacteria by the above mentioned antibiotics could bind directly to sperm head region, decrease sperm motility and induce apoptotic-like changes in stored sperm. When combined with penicillin G, polymyxin B (0.1 mg/ml; a polycationic antibiotic) blocks the negative effects of endotoxins on sperm quality and improves long-term preservability of boar semen (55).
At the legislation level, there are two reference organizations: the Office International des Epizooties (OIE) and the European Union (EU). In its International Animal Health Code (2002), the OIE regulates the criteria to be applied to semen media. This norm recommends that if a medium contains an ingredient such as milk, egg yolk or other animal protein, these should be pathogen-free or sterilized. The addition of antibiotics is permitted, provided they are declared in the corresponding international veterinary certificates. In the EU setting, Directive 90/429/CEE (1990), which regulates health policies applied to the exchange between member states and the import of boar semen, stipulates the use of a combination of antibiotics that should be efficient particularly against leptospirochaetes and mycoplasma. Its concentration should at least have an effect equivalent to the following (per ml): 500 IU penicillin, 500 IU streptomycin, 150 mg lincomycin and 300 mg spectinomycin. Replacement of streptomycin by gentamicin (0.25 mg/ml) in the latter combination of antibiotics has no negative effect on sperm quality. This norm also indicates that immediately after adding the antibiotics, the diluted semen should be kept at a temperature of at least 15°C for a minimum of 45 min (2, 12, 56).
It is worth noting that there is an increasing move towards supplying semen extenders in a liquid form (rather than as a powder which is dissolved in distilled water immediately prior to use). Since antibiotics have a limited half-life in the liquids, the manufacturers add high concentrations of antibiotics in order to retain a sufficient antibiotic activity over the shelf life of semen extender. This calls the attention towards the potential of development of resistant bacterial strains for these antibiotics. Therefore, any reduction in the use of antibiotics in semen extenders would be desirable (57, 58).
Despite colloidal centrifugation of semen reduces, to some extent, bacterial contamination of boar ejaculates, the harvesting of sperm pellets is performed in a laminar air flow bench, whereas most semen collection stations do not have access to such equipment. This raises the question about the likelihood of re-contaminating the “clean” sperm pellets with bacteria from the environment. Moreover, motile flagellated bacteria are more likely to be present after colloidal centrifugation than non-flagellated bacteria. Under normal conditions, boar semen doses are stored in sealed plastic containers with minimal amount of air. Such anaerobic conditions would also be unfavorable to bacterial growth. It should be noted, however, that neither the purification of semen nor the use of antibiotics is a substitute for good hygienic practices on boar stud farms (14, 57).
1. Principles behind anaerobic storage of semen at 15-20°C
In freshly ejaculated boar semen, pH varies between 7.2 and 7.5, and below this pH range, sperm motility and metabolism are reduced gradually (2). In sperm cells, the initial production of lactic acid from metabolism of the high glucose content of most semen extenders (e.g. BTS) causes a considerable reduction of intracellular pH to 6.2. Since diluted boar semen is placed routinely in plastic tubes with little or no access to oxygen, the accumulation of non-oxidized lactic acid causes intracellular acidosis and subsequent cessation of sperm metabolism and motility. In theory, these conditions enable porcine spermatozoa to survive storage for some days at ambient temperature (23, 25, 27, 40, 59-65). Furthermore, maintaining stored boar semen at ~ 16°C will optimize sperm quality characteristics (66).
Incubation of epididymal boar sperm at room temperature for 30 min with seminal plasma, rather than Ringer-fructose buffer, is associated with low progressive motility and retention of cytoplasmic droplets. This may be due to the presence of insufficient levels of energy substrate in the seminal plasma. However, the initial sperm motility (within 5 min) after dilution in the fructose containing buffer is oscillatory (progressive & total motility = 2-25% & 78-84%) and sperm cells acquire their motility on incubation (progressive & total motility = 64-78% & 88-92%) for 15-30 min. This has important implications in semen processing: first, dilution of semen should be done with media containing sufficient energy substrates, thus spermatozoa can initiate sperm motility to help shedding off their droplets and then enter a state of cellular anabiosis due to anaerobic conditions of semen preservation. Second, before examination of liquid-stored semen, it is necessary to incubate the samples for at least 30 min under aerobic conditions at 37°C (23, 34, 41, 67-69).
The pH of semen storage media normally use ranges from 6.8 to 7.2, but it should be taken into account that in these media, the pH does not become stable until 60-90 min from the start of dilution with water and that the different extenders show a different pattern of pH change over time (12). In addition, appropriate measures should be considered when preparing the extender by minimizing its contact with the air and thereby avoiding loss of CO2 from the buffering system of the extender (62), a finding which may explain, in part, the detrimental effects of pH rise during semen preservation on sperm motility (70).
Boar spermatozoon has an osmotic pressure of 290-300 mOsm/Kg and can tolerate a relatively wide range of osmotic pressures (240-380 mOsm/Kg). Neither sperm motility nor viability is affected by osmotic pressures in the range of 250 to 300 mOsm/Kg. However, at osmolarity below 200 mOsm/Kg, sperm motility is significantly reduced (71-73). In the commercial use, isotonic or slightly hypertonic media (300-330 mOsm/Kg) have provided better preservation of sperm fertilizing capacity than hypertonic media (12).
The ideal dilution rate of semen with pure electrolyte media ranges between 1:8 and 1:11 for sperm motility and acrosomal integrity. However, dilution titers of 1:9 to 1:11 consistently maintain the highest percentage of sperm motility (74). This is of importance during semen preparation for AI: if the calculated dilution titer lies in the normal range (≥ 1:8 and ≤ 1:11), so extend the semen at the estimated rate; If the calculated dilution titer is below the lower limit (1:8), it is recommended to dilute the semen at 1:8; and If the calculated dilution titer is above the upper limit (1:11), it is recommended to dilute the semen at 1:11. The optimum dilution titer differs for each semen medium used for sperm preservation. Initial sperm concentration in the ejaculate and characteristics of the medium, such as initial pH, buffering capacity, and osmotic pressure, can influence the optimum dilution titer during storage (74). Certainly, a 1:10 dilution of sperm-rich fraction of an ejaculate (probably averaging only 60x106sperm/ml during storage) is not comparable to a 1:10 dilution of the whole ejaculate. Furthermore, the lactic acid concentration in a stored sample of semen will be proportional to sperm concentration. Consequently, a medium with a poor buffering capacity requires a higher dilution rate than a medium with a good buffering capacity (74).
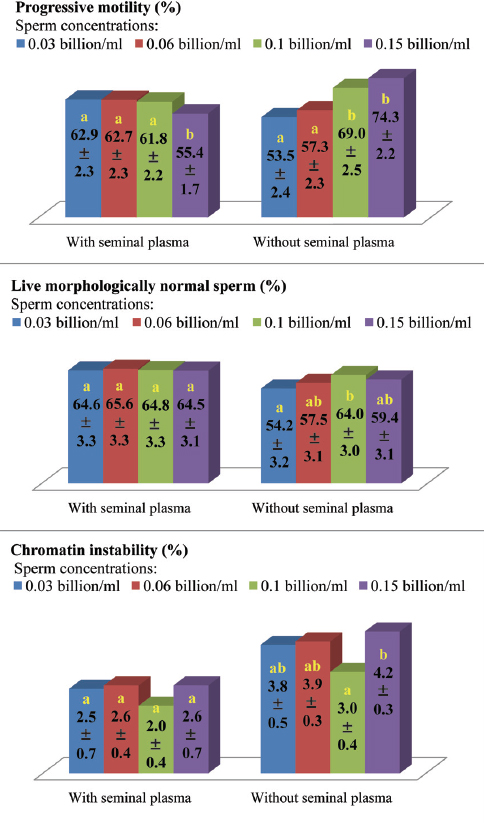
High dilution titers have negative effects on sperm viability and boar seminal plasma protects sperm integrity against this ‘dilution effect’ (75). Recent observations in our laboratory support this hypothesis (21), where significant effects were detected for the interaction between sperm concentration and presence or absence of boar seminal plasma during semen storage on sperm motility, morphology and chromatin stability (Fig. 1). Under the practical conditions, high dilution rates are likely to be a problem than low dilution rates, because keeping the entire inseminate dose in one container is less complex than the initial semen storage at a low dilution titer and then addition of the medium just before AI to provide the optimal inseminate volume (2, 74-76).
Figure 1. Influence of sperm concentration and presence or absence of seminal plasma during storage of boar semen at 17°C for 24 h in INRA-96 extender (IMV technologies, France) on sperm quality traits (21). Means (±SEM) with dissimilar letters within each semen treatment, i.e. with or without seminal plasma, are significantly different at P < 0.05-0.01.
As boar spermatozoa show a higher percentage of circular movement than those from other species, it is recommended to estimate the different forms of motility, including the progressive motility. Stored semen should be examined daily, with a value of sperm progressive motility above 60% is considered satisfactory (2). The presence of cytoplasmic droplet (proximal or distal) is a serious morphologic defect, which is of particular importance when semen stored for long-term is used for AI and may be associated with lowered resistance of spermatozoa against in vitro aging (77). The total percentage of cytoplasmic droplets in ejaculates used for AI should not exceed 15%, especially when stored semen is used. The percentage of other morphological alterations should not exceed 20% (2, 77). Strictly, liquid-stored semen used for AI should contain > 60% progressively motile sperm and ≥ 85 % morphologically normal spermatozoa.
The fertility of liquid-stored boar semen depends mainly on the following factors: initial ejaculate quality and frequency of semen collection, extender type, storage temperature and duration, sperm number per AI dose, insemination volume, number and timing of inseminations relative to ovulation at each estrus as well as the site of semen deposition in the female genital tract (16, 39, 45, 77-92). In addition, a number of factors can also influence the fertility levels in pig farms, including boar breed and age, parity and lactation length, weaning-to-estrus interval, season, stress, nutritional status and inseminator skills (80, 85, 86, 88, 91, 93-100). In the authors’ experience, these factors should be taken into account before designing the clinical studies because they may have a confounding influence on the outcome of field-fertility trials. Therefore, the readers are directed to the above stated references for expanding their knowledge regarding the factors which affect the farrowing rate and sow prolificacy after AI with preserved porcine semen.
In the commercial situation, the principal aim of semen storage is to maintain spermatozoa in a viable state for a variable period of time in order to fertilize a high proportion of ova with a minimum sperm dose and at minimal cost and health risk (2). This aim has not been achieved by BTS extender when storage time exceeds 3 days. However, doubling of the sperm dosage to 6 billion on the fourth day of storage along with the use of boars with good semen quality can prevent the decline in semen fertility (2, 81). Unfortunately, this option is not usually practicable for the commercial use of liquid-stored boar semen. It is our opinion that the higher cost of the long-term extender, such as Androhep® or X-cell®, is acceptable because of the higher fertility and prolificacy levels after semen storage for 4-5 days and the possibility of flexible semen handling (2, 77, 82, 87, 90).
2. Processing steps for liquid storage of semen using short-term (modified BTS) and long-term (MOPS) extenders in our laboratory (101)
► Preparation of modified BTS extender: 3.7 g D-(+)-glucose anhydrous, 0.6 g Na-citrate tri-basic 2H2O, 0.125 g EDTA-2Na, 0.125 g NaHCO3, 0.075 g KCl, sterile-reverse osmosis water up to 106.255 ml, 0.5 ml of 5% gentamicin sulfate solution, and 20 μl of citric acid-Hepes solution (0.5245 g citric acid H2O + 0.5958 g Hepes + 2.5 ml water). The medium (~ pH 7.2 and 310 mOsm/Kg) is then sterilized through a 0.22-μm filter.
► Preparation of MOPS extender: 1.2566 g D-(+)-glucose H2O, 1.733 g Na-citrate tri-basic 2H2O, 0.2623 g EDTA-2Na, 0.1319 g NaHCO3, 0.4918 g Mops, 0.5 g bovine serum albumin, 0.06 g penicillin G Na (≥ 1477 U/mg), 0.01 g neomycin sulfate, 0.01 g kanamycin sulfate and sterile-reverse osmosis water up to 100 ml; adjustment of medium pH (with 1M solution of citric acid H2O) at 6.7-6.8 and osmolarity at 290-300 mOsm/Kg. The extender is then sterilized through a 0.22-μm filter.
► Warm the medium to 37°C in a water bath.
► Collect a whole ejaculate in a pre-warmed container using the “gloved-hand” technique.
► Incubate the gel-free semen at 37°C and determine ejaculate volume, sperm concentration and progressive motility.
► Calculate the amount of extender required for dilution of the ejaculate based on:
* Semen dilution titers (semen: medium) = 1:1-1:11 (optimum 1:7.5).
* Insemination volume = 80-100 ml.
* Insemination dose = 3x109 spermatozoa.
* Post-dilution sperm progressive motility ≥ 75%
* Post-dilution morphologically normal sperm ≥ 85%
► Add the medium to the semen in a stepwise manner and incubate the diluted ejaculate at 37°C for 5-10 min.
► Cool the semen to the required temperature (20-16°C) at a rate of – 0.10°C/min.
► Package the cooled semen. The air-filled spaces above the semen should not be larger than 4.5-10% of the tube’s or bag’s capacity.
► Store the insemination doses at 16-20°C (optimum 18°C).
► Semen should not be used for AI before 45 min of the dilution process.
► Semen doses should be rotated at least twice a day during the whole storage period.
► Under the above recommendations, semen fertility and prolificacy can be preserved up to 72 and 120 h in modified BTS and MOPS extenders, respectively.
1. Theoretical bases behind cryopreservation of boar semen
Boar semen differs in several aspects from that of human and other domestic animals. It is produced in large volumes and is extremely vulnerable to cold shock or sudden cooling immediately after collection (30, 34, 102, 103). Boar spermatozoa are also susceptible to warm shock when cooled semen (5°C) is rapidly diluted at 25-37°C (104, 105). Prolonged exposure of spermatozoa to seminal plasma is detrimental to their motility and morphology during cooling and holding of the semen at 5°C (Fig. 2) (21). These characteristics require special considerations in the design of freezing protocols for porcine semen.
Figure 2. Influence of extender and presence or absence of seminal plasma during storage of boar semen at 5°C for 24 h on sperm quality characteristics (21). Means (±SEM) with dissimilar letters are significantly different at P < 0.02-0.0.001.
Successful freezing of boar semen relies on understanding of the factors and their interactions which influence the capacity of spermatozoa to survive freezing and thawing. These factors can be classified into two categories: internal or fixed factors, such as the inherent characteristics of spermatozoa as well as the differences between breeds, boars and ejaculates, and external factors, such as composition of storage media, rates of dilution and cooling, type and concentration of cryoprotective agents, equilibration, and method of freezing and thawing of semen (2, 106).
Cooling phase
Boar spermatozoa acquire a resistance to cold shock simply by incubation at 17-30°C for several hours (36-38, 107-111). Various factors affect the development of sperm resistance to cold shock. First, seminal plasma of whole ejaculate: acrosomal membranes of epididymal spermatozoa are more resistant to cold shock damage than those of the ejaculated ones (30, 112). This is attributed to sperm acquisition of a proteinous coat derived from secretions of the seminal vesicle. These proteins are strongly basic, very difficult to dislodge from sperm surface and are capable of increasing permeability of cellular membranes (34). This may explain why sperm-rich fraction, which contains only a low proportion of seminal plasma, develops a more resistance to cold shock than the whole ejaculate (36). Within the sperm-rich fraction, the first 10 ml (P1) are more tolerant to cold stress than the rest of sperm-rich fraction and the post-sperm fraction (P2). Such tolerance is influenced by seminal plasma of the portions because seminal plasma of P1 increases the resistance of P2-spermatozoa to P1 levels (113).
Second, intrinsic cellular property: washed sperm suspensions have higher percentages of normal acrosome and motility when cold-shocked after 5-h incubation than after 1- or 3-h incubation, indicating that the ability of spermatozoa to develop cold shock resistance during in vitro incubation is an inherent trait within these cells (37). Supporting this hypothesis, when epididymal spermatozoa are extended (1:3) in a Tris-lactose medium and cold-shocked to 0°C after 1- and 5-h incubation, percent sperm with normal acrosome is 4% and 30%, respectively (37).
Third, incubation period and temperature: increasing the incubation time is associated with increasing sperm resistance against cold shock (progressive motility and acrosomal integrity after 2 and 4.5 h, respectively) (108). Sperm acrosome and motility acquire a more cold shock resistance when cold shock was done after 2.5-, 3- and 5-h incubations than after 0.5- and 1-h incubations (36, 37, 108). No studies were conducted regarding the effect of incubation temperature on cold shock resistance of boar sperm. However, maintaining of diluted semen at ~16°C overnight improves quality of frozen spermatozoa (66). On the other hand, although extending the holding period of semen from 3 to 24 h at 15°C has no significant influence on freezability and pregnancy rate of frozen spermatozoa, it reduces the potential litter size (114).
Fourth, sperm dilution and seminal plasma of sperm-rich fraction: increasing the dilution rate of sperm-rich fraction from 1:2 to 1:6 or 1:10 interferes with the development of cold shock resistance by boar sperm during in vitro incubation. Cold shock affects acrosome structure and sperm motility more adversely as the sperm concentration decreases from 278 to 56 x106/ml, even though the seminal plasma and spermatozoa make up to 50% of the final volume of the diluted sample, i.e. at a fixed seminal plasma concentration. It seems that dilution per se increases susceptibility of boar sperm to cold shock because further dilution of 3- and 5-h incubated semen from a rate of 1:2 to rates of 1:6-1:10 decreases sperm motility and acrosomal integrity compared with incubated-diluted semen at a rate of 1:2 (37). However, seminal plasma per se also provides boar sperm with some protection against cold shock damage after 3- and 5-h incubation. It appears that seminal plasma interacts with spermatozoa during incubation to produce the increased resistance to cold shock. In agreement with this opinion, incubation (10 min at room temperature) of cauda epididymal spermatozoa in seminal plasma from intact and vasectomized boars increases their resistance to cold shock by 60-80% (112). Therefore, the detrimental effect of excessive semen dilution on resistance of boar spermatozoa (sperm-rich fraction) to cold shock may stem from a direct adverse effect of dilution on sperm cells, excessive dilution of seminal plasma constituents and a direct interference with incubation-induced development of cold shock resistance. Interestingly, recent studies demonstrate the beneficial effects of low levels of boar seminal plasma on quality and fertility of cooled (5°C) and frozen-thawed semen (113, 115-120).
Fifth, incubation medium: sperm incubation medium has a significant effect on the development of tolerance to cold stress (36, 121). When the sperm-rich fraction is cold-shocked after 2-h incubation, motility in media with pH 7.3-8.3 are higher than in those with pH 5.9-6.6. After 4.5-h incubation in media with pH range of 5.9-8.3, acrosomal integrity of cold-shocked sperm is higher than that after 1-h incubation. However, sperm acrosomes are more resistant to cold shock after 7-h incubation in media with pH 5.9-7.3 than in media with pH 8-8.3 (108).
Disaccharides (lactose and sucrose) and trisaccharides (raffinose) maintain acrosomal integrity better than monosaccharides (glucose and fructose) after cold shock of 5-h incubated spermatozoa. Fructose maintains acrosomal integrity better than glucose. Sugars do not influence sperm motility after cold shock of incubated semen. Varying levels (0.082-0.246 M) of Tris in the media have no effect on the development of cold shock resistance during semen incubation for 1-5 h (108). Egg yolk (5-30%) does not protect boar sperm motility and acrosome against cold shock after 1 and 5-h incubation. Sperm acrosomes are more cold shock resistant after 5-h incubation in the absence rather than in the presence of egg yolk. Therefore, egg yolk (20-30%) interferes with development of cold shock resistance (108). In the absence of seminal plasma of sperm-rich fraction, milk casein (2-4%; in Tris-lactose medium; ~ 310x106 sperm/ml) protects sperm acrosome, but not motility, against cold shock damage. However, in the presence of seminal plasma, casein does not provide such protection to sperm acrosome (37).
Boar spermatozoa differ markedly in their response to cold shock compared with bull and ram spermatozoa (2). Chicken egg yolk, lecithin (phosphatidylcholine), cow milk, casein, albumin and globulin have been reported to alleviate cold stress in equine, bovine, ovine and caprine spermatozoa (122-126). On the contrary, whole egg yolk, albumin, globulin, phosphatidylcholine and phosphatidylethanolamine do not protect porcine sperm against cold shock (37, 127). Only phosphatidylserine is able to safeguard boar sperm against cold shock (127). Orvus Es Paste (OEP; also known as Equex STM; currently distributed by Nova Chemical, USA and Minitüb, Germany), a synthetic detergent based on sodium and triethanolamine lauryl sulfate, modifies egg yolk constituents and renders egg yolk extenders favorable for low-temperature storage of boar spermatozoa (128-131).
Butylated hydroxytoluene (BHT), a lipophilic chain-breaking antioxidant, has been detected (alone or in combination with egg yolk) to protect spermatozoa from the damaging effects of cooling and warming of boar semen (19, 104, 105, 132-135). However, BHT alone is effective only when spermatozoa are cooled to 5°C at a slow rate (133). In our lab, despite we demonstrated that BHT could be an alternative of egg yolk for cooling of goat semen (136, 137), we did not observe a similar function for BHT on motility and acrosomal integrity of cooled (5°C) boar semen (unpublished data), a finding which is in agreement with results of earlier studies on cold-shocked spermatozoa (138, 139). However, a combination of BHT and egg yolk improves boar sperm preservability (135). These observations suggest that it may not be realistic to expect a single compound to stabilize the entire boar sperm plasma membrane during cooling or freezing, but those combinations of lipids and other membrane-stabilizing agents may be required to adequately protect all membrane domains (139).
The low-density lipoprotein (LDL) fraction of avian egg yolk (140-144) and soybean lecithin (145) have also been shown to preserve the structural and functional integrity of cooled (4°C) and frozen-thawed boar spermatozoa. However, we observed in our lab that soybean lecithin was not as efficient as egg yolk in supporting motility of frozen-thawed boar spermatozoa (unpublished data). Skim milk powder alone or in combination with egg yolk has also been shown to protect boar spermatozoa during cold storage (130, 146). Treatment of porcine spermatozoa with cholesterol-loaded cyclodextrins (CLC) increases their tolerance to cold stress during low-temperature processing of semen samples (18, 31-33).
In short, evidences from empirical studies show that skim milk, casein (BF3 extender), BHT in combination with egg yolk, OEP-supplemented egg yolk (TEST and BF5 extenders), phosphatidylserine, LDL of egg yolk and CLC can contribute to the protection of boar sperm during the initial cooling (5°C) phase of semen freezing (32, 127, 130, 131, 135, 146).
Freezing phase
Many cryoprotective agents have been tested, but none have proved better for preserving boar spermatozoa than glycerol (103). However, porcine sperm cells show a greater sensitivity than those of other domestic animals to glycerol levels adequate for optimum freezing (147). The concentration of glycerol required for maximum survival of spermatozoa is determined by several considerations; one of them is the freezing rate. When static liquid nitrogen vapor and dry ice are used for freezing, the volume and geometry of the semen sample influence the rate of freezing and glycerol concentration used (2). Nevertheless, it is generally accepted that only relatively low concentrations of glycerol may be used to obtain a satisfactory post-thawing sperm survival (128, 131). Glycerolated media are usually added when the partially diluted semen has been cooled to 5°C. Although the composition of diluents varies, most of them are characterized by low glycerol levels (2.5-4%) and a short equilibration period (2, 130).
Revisiting the semen-glycerol equilibration time for semen packaged in 0.5-ml straws and protected with various glycerol levels (0-6%), indicates that longer times of exposure to 5°C are beneficial, with a 4-h equilibration producing the highest value of motile sperm with normal acrosome. As semen with no glycerol benefits from the exposure to 5°C similar to glycerolated semen, it seems likely that the holding time at 5°C is solely renders spermatozoa less susceptible to the freezing damage (148).
The interaction between glycerol concentration and cooling velocity has been recognized as an inverted U-shape survival curve for boar spermatozoa frozen over a range of cooling rates from 1°C to 1500°C/min (147, 149). The optimum cooling rate shifts to higher values with the decreasing glycerol concentration. It is also shown that the optimum glycerol concentration varies with the parameter investigated: for sperm motility, the best glycerol concentrations are 3-4% whereas for acrosomal integrity, the optimum concentration is 0-1%. This is attributed to the complicated structure of the spermatozoon which comprises several geometries and different regions. The optimum conditions for one part of the cell may be unsuitable for other regions, or the damage to some parts may have greater consequences than to the others (103). Thus, for preservation of both sperm traits, it is recommended a compromise with 3% glycerol and freezing rate of 30°C/min (149). It is worth noting that the gradual decrease in motility and acrosome integrity of boar sperm cells as the glycerol concentration exceeds 6% may be due to the chemical toxicity of glycerol, or to the osmotic shock by rapid dilution of the glycerol at thawing (103,105).
Warming phase
The rate of thawing of frozen semen through the critical temperature range is an important factor affecting survival of spermatozoa (150). In boar semen frozen at the optimum rate (e.g. 30°C/min) in 0.5-ml straws (149), sperm motility and acrosome integrity are improved with increasing the thawing rate. This is consistent with the well known ‘two-factor’ hypothesis that cryoinjury after rapid freezing is caused predominantly by the re-growth of minute ice crystals during slow thawing and that a high warming velocity is essential for sperm cryosurvival (151, 152), although no convincing evidence of intracellular ice formation has ever been obtained for sperm cells (150). In fact, recent evidence shows that sperm damage at rapid rates of cooling is a result of an osmotic imbalance encountered during thawing, not intracellular ice formation (150). Three findings may support this evidence: (a) the effect of thawing rate on the acrosome integrity is influenced by the osmotic stress induced by the glycerol concentration, higher concentrations being more damaging to acrosomal integrity (147), (b) rapid warming of cooled-glycerolized (1%) boar semen to 37°C causes a significant damage in sperm acrosome, which can be minimized by pre-treatment of spermatozoa with the antioxidant, BHT (105), and (c) post-thaw dilution of frozen boar sperm causes a significant reduction in their quality traits (Tables 1-3) (21), probably due to a hyperosmotic stress imposed by rapid removal of glycerol from thawed spermatozoa (113, 153) and/or an oxidative stress caused by removal of seminal plasma constituents during semen processing (75). Indeed, recent studies demonstrate that exposure of sperm cells to anisosmotic stress stimulates production of reactive oxygen species (154) and post-thaw supplementation of frozen boar semen with seminal plasma improves sperm quality and fertility (115, 120).
In brief, according to the procedures developed quasiempirically over the past decades, effective freezing of boar sperm requires the optimum combination of three main variables: glycerol concentration, cooling and warming velocities. Considering the maintenance of sperm motility and acrosome integrity, the optimum represents a compromise: 3% glycerol, a freezing rate (in the range between 0°C and – 50°C) of 30°C/min and a thawing rate of 1200°C/min (2).
2. Processing steps for frozen storage of semen in our laboratory (21)
► Preparation of semen processing media:
* Warm the holding medium to 37°C. The medium is composed of 1.9636 g Tris, 0.9567 g citric acid anhydrous, 4.9154 g α-lactose H2O, 0.2960 g D-(–)-fructose, 0.0395 g KCl, 0.0991 g EDTA-2Na, 0.01 g kanamycin sulfate and sterile-reverse osmosis water up to 100 ml. The medium (pH 7.3-7.4 and 330 mOsm/Kg) is then sterilized through a 0.22-μm filter.
* Cool the cooling medium to 18°C. The medium consists of 7.5 ml chicken egg yolk, 92.5 ml INRA-96 extender and 0.1 g EDTA-2Na. INRA-96 is a ready-to-use sterile extender (pH 6.9; 307 mOsm/Kg) containing a solution of Hank’s salts, glucose and lactose, buffered by Hepes and supplemented with native phosphocaseinate (27 g/L), penicillin (50 IU/ml), gentamicin (50 μg/ml) and amphotericin B (0.25 μg/ml) (IMV technologies).
* Cool the freezing medium to 5°C. The medium consists of 7.5 ml chicken egg yolk, 9 ml glycerol (> 99%), 83.5 ml INRA-96 extender and 0.1 g EDTA-2Na.
► Semen collection:
* Collect good quality samples (sperm-rich fractions) in a pre-warmed container from boars with known sperm freezability ‘good freezers’.
* Incubate the semen at 37°C and determine semen volume, sperm progressive motility and concentration using a pre-calibrated photometer.
► Primary holding and cooling (development of sperm resistance to cold shock):
* Extend the semen with the holding medium at a dilution titre (semen: medium) of 1:1 to 1:2.
* Cool the semen from 37°C to 18°C at a rate of – 0.10°C/min (~ 3.2 h).
* Estimate sperm cell concentration using a Neubauer hemocytometer chamber.
► Centrifuge the semen at 2400 x g for 3 min (18°C) for removal of seminal plasma and concentrating of spermatozoa.
► Secondary cooling (cooling of spermatozoa to 5°C):
* After centrifugation and removal of seminal plasma, re-suspend sperm pellet in the cooling medium at 18°C to a concentration of 1.5x109sperm/ml.
* Cool the semen from 18°C to 15°C at a rate of – 0.1°C/min and then from 15°C to 5°C at a rate of – 0.067°C/min (~ 3 h).
► Secondary holding, glycerolization and equilibration (development of sperm resistance to freezing damage):
* After cooling, hold the semen at 5°C for 1 h.
* Add the freezing medium to the cooled semen at a ratio of 1 part of the medium to 2 parts of cooled semen to achieve concentrations of 1.0x109sperm/ml and 3% glycerol. The addition of freezing medium should be done over a 20-min period in 4 fractions at 5-min intervals.
* Equilibrate the glycerolized semen at 5°C for 1 h before freezing.
► Packaging of equilibrated semen:
* Fill 0.50-ml straws pre-cooled to 5°C with semen (400-450 μl/straw).
* Seal the straws and place them horizontally onto pre-cooled (5°C) freezing racks.
► Two methods are recommended for freezing of semen:
* Horizontal exposure of straws to static liquid nitrogen (LN2) vapor: maintain the straws for 20 min at 3 cm above the level of LN2 and then plunge them into LN2.
* Controlled-rate freezing: transfer the straws horizontally in the racks to a programmable freezer pre-cooled to 5°C. Freeze the straws using one of the following freezing curves:
(a) From +5 to – 4.5°C at –1°C/min, 60 s at – 4.5°C, from – 4.5°C to – 180°C at –30°C/min and then plunge the straws into LN2.
(b) From +5 to – 5°C at – 6°C/min, from – 5°C to – 80°C at – 40°C/min, 30 s at – 80°C, from – 80°C to – 150o C at – 70°C/min and then plunge the straws into LN2.
► Thawing of frozen semen:
* Warm the modified BTS extender to 37°C.
* Individually immerse the frozen straws in a circulating water bath at 50°C for 20 s.
* Incubate the frozen-thawed semen at 37°C for 5-10 min.
* Extended the semen to 0.05-0.3x109sperm/ml with BTS. In order to minimize the osmotic stress on spermatozoa, post-thaw dilution of frozen semen should be done gradually over a 20-min period in 4 fractions at 5-min intervals.
1. Colloidal purification of porcine semen
Concepts behind isopycnic purification of semen samples
In AI programs, some boars of high genetic value may have ejaculates of low quality traits and poor preservability. These ejaculates are always discarded before or after semen processing leading to economic losses in view of decreasing availability of insemination doses from genetically superior sires. The presence of abnormal or dead sperm, bacteria and leukocytes in semen samples has been suggested to have a detrimental effect on the fertilizing potential of AI doses (14, 42, 48-51, 155, 156).
Similarly, the occurrence of bacterial endotoxins and mycotoxins in boar ejaculates may be responsible for a rapid deterioration in sperm quality traits during liquid storage of semen (55, 157-162). In addition, studies on hygienic aspects of semen storage for AI indicate that supplementation of extenders with antibiotics is not enough to eliminate the detrimental influence of bacteria on sperm motility, viability and fertility (52, 163, 164). There is also a potential risk of transmitting pathogenic microorganisms through AI with stored semen (47, 52, 163-165).
Consequently, purification of porcine semen aims at: (a) improving quality and storageability of AI doses by selecting spermatozoa with competent structural and functional integrity as well as by reducing the concentration of bacteria and toxins which may contaminate boar ejaculates (55, 158, 166-169), (b) raising the biosecurity standard of AI programs by removing the potential pathogenic bacteria and viruses from semen doses (57, 58, 164,170), and (c) teleologically, minimizing development of antibiotic-resistant strains of bacteria through decreasing the use of antibiotics in storage extenders (57, 164).
Ejaculated spermatozoa are characterized by a pronounced heterogeneity in morphology, motility, viability and genomic integrity (172). When semen samples are centrifuged through a column of colloidal silica particles, cells move to a point that matches their own density–the isopycnic point, and by altering the centrifugation force and time along with the physical properties of the colloid, the good quality spermatozoa are allowed to pellet in the bottom of the tube (58, 173-177). Several intrinsic properties of colloidal silica make it ideal for preparing density gradients for selecting sperm cells. First, as a mineral substance, it has no osmotic effects when added to the culture medium; second, it allows high-density (high specific gravity) media to be prepared, which is important because normal, mature mammalian spermatozoa are dense cells; and third being a colloid rather than a solution, it has a low viscosity, and thus it does not retard sedimentation of sperm cells (174). Some 20 years ago, silane-coated silica colloid became available that could be sterilized by autoclaving and it had a low level of endotoxins (58). Silane-coated silica colloid has the advantage of being stable for long periods in salt solutions, and hence permitting standardized formulations to be sold commercially (58). Two products of ready-to-use silane-coated silica colloid are currently marketed for boar semen, namely PociPure™ (Nidacon International, Mölndal, Sweden) and Androcoll™ -P (Swedish University for Agricultural Sciences, Uppsala, Sweden) (57, 167, 170).
Procedures used in our lab for single layer centrifugation of semen over PociPure™
► Equipments and media required:
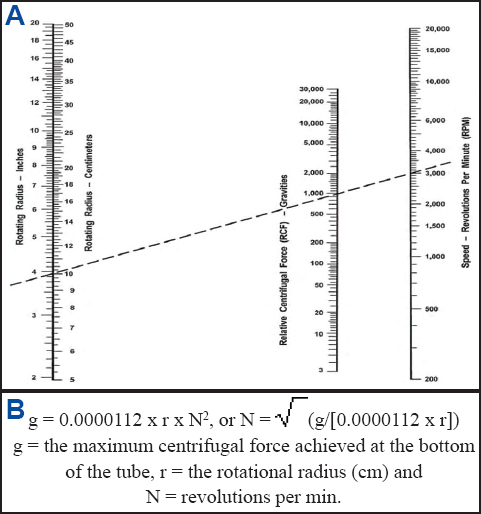
* Table-top centrifuge with a swinging-bucket rotor (Fig. 3).
Figure 3. (A) Table-top centrifuge with a swinging-bucket rotor that accommodates 50-ml centrifuge tubes and can generate a relative centrifugal force of 300-1000 times gravity. (B) Determining the rotating radius of a centrifuge by measuring the distance from the center of the rotor to the bottom of a bucket in “swinging” position. In the centrifuge shown in the image, the rotating radius is 17 cm. The length of the rotating radius is plotted against the revolutions per minute (RPM) on a relative centrifugal force nomograph chart to determine the force times gravity (g-force) as shown in Fig. 4; adapted from Vanderwall (178).
* Disposable sterile plastic conical bottom 50-ml centrifuge tubes.
* Disposable sterile “serologic” pipettes (5 and 10 ml) equipped with a manual pipettor (e.g. bulb or thumb-wheel style) for aspirating supernatant fluid.
* Prepare and warm the modified BTS (antibiotic-free) medium to 37°C.
* Prepare and incubate the MOPS extender at room temperature (RT, 22°C). The concentration of antibiotics in the extender can be decreased to 20% of its original concentrations described above. Also, incubate PociPure™ colloid at RT.
► Semen collection and evaluation:
* Collect gel-free whole ejaculates or sperm-rich fractions in a pre-warmed container using the ‘gloved hand’ technique and incubate the semen at 37°C.
* Determine semen volume and sperm concentration, e.g. whole ejaculate: 200 ml & 0.2x109sperm/ml; sperm-rich fraction: 25-50 ml & ≥ 0.8-1x109sperm/ml.
► Semen dilution:
* Extend the entire semen sample with isothermic BTS to 0.1x109sperm/ml.
* Incubate the diluted semen at RT for 15 min.
► Centrifugation of diluted semen:
* Determine the centrifugation speed using a nomograph chart or a formula (Fig. 4).
Figure 4. (A) Relative centrifugal force (RCF) nomograph chart for calculating the force times gravity (g-force) from a known rotating radius and revolutions per minute (RPM). To determine the g-force, place a straightedge on the nomograph chart connecting the known RPM and known rotating radius (in inches or centimeters) of the centrifuge. The point at which the straightedge intersects the RCF axis is the g-force. The dashed line represents an example of an RPM of 3000 and rotating radius of 10 cm that produces a g-force of 1000; adapted from Vanderwall (178). (B) A formula describes the relationship between rotation speed, rotational radius and centrifugal force (174).
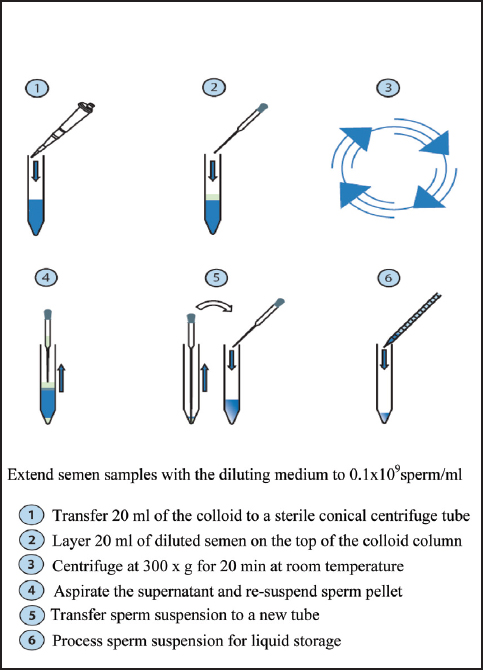
* Transfer 20 ml of PorciPure colloid to a centrifuge tube.
* Layer 20 ml of diluted semen on the top of the colloid column, resulting in a total volume of 40 ml (semen to colloid ratio = 1:1).
* Centrifuge the semen/colloid column at 300 x g for 20 min at RT.
* Aspirate the supernatant and re-suspend sperm pellet in an initial volume of MOPS extender at RT. Examine sperm motility and determine sperm concentration.
* Process the purified sperm suspension (Fig. 5) for liquid storage as described above.
Figure 5. A protocol of porcine sperm selection (PorciPure™-Nidacon)
2. Modulation of the uterine defense mechanism
Theoretical background
After AI, the number of spermatozoa in the female genital tract of the sow decreases to 5-10% within a few hours. Up to 50% of the spermatozoa are lost due to the back flow of semen in the first few hours after AI. Another important cause of the rapid decrease of the number of spermatozoa in the genital tract is phagocytosis of spermatozoa by polymorphonuclear neutrophils (PMNs). These cells migrate into the uterine lumen directly after AI and within 2 h their number reaches the same as the number of spermatozoa. Teleologically, PMNs recruitment and phagocytosis of spermatozoa can be considered as a normal physiological response to clean the genital tract in preparation for reception of the embryos. Phagocytosis of spermatozoa has also been suggested as a mechanism for preferential elimination of senescent or dead spermatozoa, or spermatozoa with a decreased fertilizing potential (179, 180).
On the other hand, treatment of boar semen to induce capacitation in vitro substantially reduces sperm phagocytosis (179). The uterine environment in vivo also provides conditions for capacitation. Spermatozoa can be fully capacitated in the uterus without ascending to the oviducts. Therefore, the reduction in phagocytosis observed after treatment to induce capacitation in vitro may also occur in vivo in the uterus. Spermatozoa may acquire a protection against phagocytosis while in the uterus, which may increase their chance to reach the isthmus and participate in the fertilization (179). In agreement with the above hypothesis, a recent investigation demonstrates that litter size increases in proportion to the number of capacitated sperm (assessed by chlortetracycline assay) in liquid-stored boar semen (181).
Caffeine, a member of the methylxanthine family, has been found to increase concentrations of intracellular cyclic adenosine monophosphate (cAMP) by inhibition of cAMP-phosphodiesterase in spermatozoa and immune cells (34, 182, 183). As a consequence, in vitroaddition of caffeine to boar semen stimulates sperm capacitation and kinematic activity, along with shedding of cytoplasmic droplets from sperm cells (69, 184-186). Moreover, it is believed that caffeine has a potent immunomodulatory effect as evidenced by its ability to decrease the chemotactic and phagocytic activities of blood PMNs when co-incubated in vitro with boar spermatozoa (187, 188).
In vivo, Matthijs et al. (180) suggests the following scenario after AI with liquid-stored boar semen: shortly after insemination, spermatozoa are still bathed in the BTS extender containing EDTA, which effectively blocks phagocytosis and capacitation as well. Most of the liquid flows back and is voided through the vulva, and about 35-45% of sperm are lost with it. Spermatozoa that are not lost with the backflow apparently have made the passage to the endometrial mucus and are thus retained. This new environment presents the conditions that allow phagocytosis, resulting in phagocytosis and digestion of large numbers of spermatozoa. In addition, these conditions allow the onset of capacitation rendering spermatozoa less vulnerable to phagocytes. Supplementation of BTS medium with caffeine and CaCl2 advances or accelerates these processes and reduces the recruitment of PMNs to the uterine lumen, leading to increasing the number of spermatozoa colonizing the oviducts, and thereby improving the probability of fertilization to occur.
Besides the above stated scenario, the procedures of porcine semen storage which involve dilution and/or removal of seminal plasma constituents, as well as the use of BTS extender and egg yolk, can exaggerate the inflammatory response of the endometrium after AI, resulting in increasing of the chemotactic and phagocytic activities of uterine PMNs (188-190). Actually, caffeine alleviates this post-insemination inflammatory reactivity, increases numbers of spermatozoa colonizing the female genital tract, and in turn improves the pregnancy and farrowing rates of gilts and sows (186, 191-193).
Procedures of caffeination of semen doses in our lab
► Preparation of the caffeination medium: 0.5652 g Tris, 3.0243 g D-(–)-fructose, 0.69 g Na-citrate tri-basic 2H2O, 0.24 g NaHCO3, 2.5 g caffeine, 0.01 g CaCl2.2 H2O and sterile-reverse osmosis water up to 100 ml; adjustment of medium pH (with 1N HCl) at 7.4-7.5. The medium is then sterilized through a 0.22-μm filter.
► Characteristics of the medium:
It is a concentrated sterile solution that, if added to liquid-stored or frozen-thawed semen just before insemination, it increases the pregnancy and farrowing rates by ~15-20 points. It can be used to maximize fertility of semen in the following cases:
* AI with reduced sperm dose (1.5-2 x109 sperm), which allows maximum utilization of semen from genetically superior sires.
* AI of sows which come early or late in post-weaning estrus.
* AI with semen having suboptimal quality spermatozoa such as frozen-thawed semen, sex-sorted semen and liquid semen that has been stored in a short-term extender > 2 days or in a long-term medium > 4-5 days.
* AI with semen obtained from young (< 1 years old) and aged (> 5 years old) boars.
* AI with liquid-stored semen during the summer season.
* AI of superovulated sows in the embryo transfer programs.
► Application of the medium:
* Warm up the medium to 37°C and transfer the required amount to the insemination bottle by a sterile 1- or 2-ml syringe. Shake gently the bottle to ensure complete mixing of the semen with the medium and then inseminate. To achieve optimum results, treated semen should be used within 1-15 min after addition of the medium.
* The medium should be added to liquid-stored or frozen-thawed semen at a rate of 10 μl medium to 1 ml diluted semen. Examples are given in the following table:
3. Site of semen deposition in the reproductive tract during AI
Intracervical AI
Liquid-stored semen
The insemination procedure of standard cervical AI deposits the semen dose within the posterior portion of the cervical canal (~ 15 cm deep into the cervix) by means of a catheter that engages the folds of the cervix (Fig. 6). In order to achieve systematic high fertility rates among pig farms, the protocol recommends deposition of 3x109 spermatozoa in a large volume (80-100 ml), two or three times during the estrus period (194). It is worth noting that while unsatisfactory results have resulted when AI dose is reduced below 2.5x109 spermatozoa, well-managed herds achieve high fertility rates with only 2x109 spermatozoa per insemination. Moreover, even 1x109 spermatozoa per dose can give good fertility provided that the timing of AI is optimal and no backflow of semen during insemination occurs (194). The optimal time for AI is in the interval from 28 h before to 4 h after ovulation. However, the highest fertility can be seen when liquid-stored semen is placed in the reproductive tract within 12-24 h before ovulation, regardless of sperm number delivered (11). Fixed-time AI (single or double) is not usual when liquid semen is used. Currently, sows are bred every 24 h from the start of the estrus until the standing reflex is no longer detected, resulting in > 2.5 average number of inseminations per estrus (11).
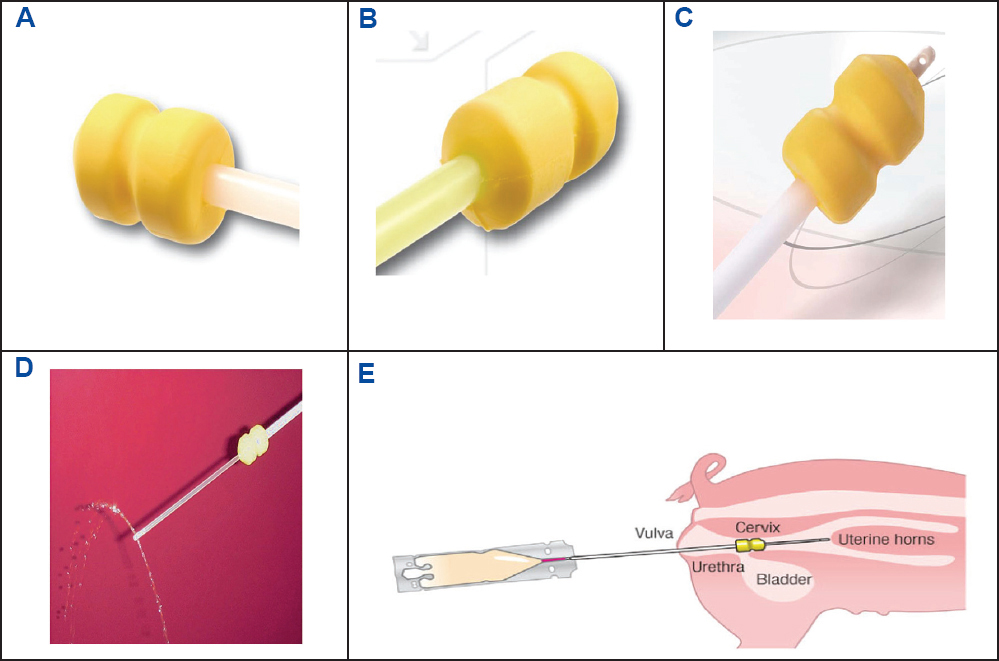
Figure 6. The polyurethane sponge tip of the conventional catheter (A) and the smaller tapered tip of the gilt catheter (B) for intracervical AI. The tip of the intrauterine catheter (C) having two lateral orifices with semen emerging (D) during AI (E); adapted from IMV technologies (France) and Watson and Behan (89).
Cryopreserved semen
During the last decade, optimal fertility results with frozen semen have been reported if sows or gilts are inseminated intracervicallly 2-3 times in the natural or induced estrus with 5-6x109 spermatozoa per AI dose (~ 2-3 x109 motile sperm per dose) (11, 199). The biological reason of this large number of sperm per AI dose may be related to weakness of frozen-thawed spermatozoa, which are especially vulnerable to loss of their functional competence during the transit through the long reproductive tract of the female pig (11). In USA, a feasibility study for freezing cost per ejaculate is estimated at $275 per ejaculate or $11 per dose of frozen-thawed semen at standard doses of 5x109 total frozen sperm. This cost estimate does not include genetic value, fixed equipment costs, depreciation, or variable lab space fees (199).
Transcervical AI
Despite the high number of sperm delivered via cervical AI, most of them do not participate in the fertilization process because they are rapidly evacuated from the reproductive tract by backflow during or immediately after AI (30-40% of the total number of spermatozoa deposited into the cervical canal), trapped and die in the cervical folds (5-10%) and phagocytosed in the uterus (up to 60%) (11). Therefore, several different instruments have been developed to transverse the cervix and deposit the semen either in the uterine body “posterior horn” (intrauterine insemination; IUI) (89, 195) or in the uterine horn (deep intrauterine insemination; DIUI) (196, 197).
Nevertheless, careful attention should be paid to proper transcervial AI technology to minimize the incidence of cervical and uterine damage which negatively influences the reproductive longevity of sows and gilts in the farm (194, 198).
IUI with liquid-stored semen
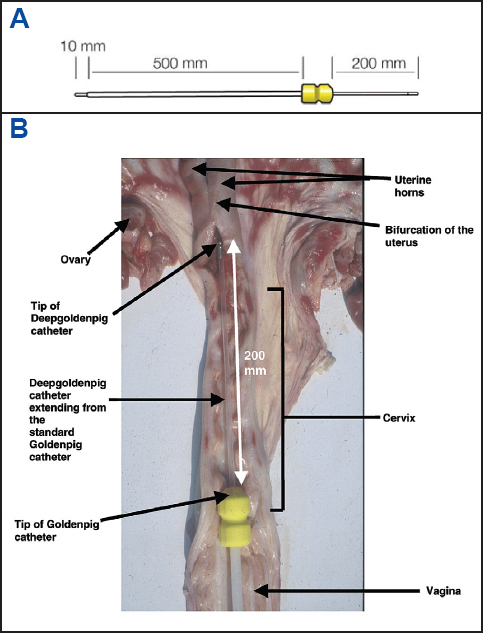
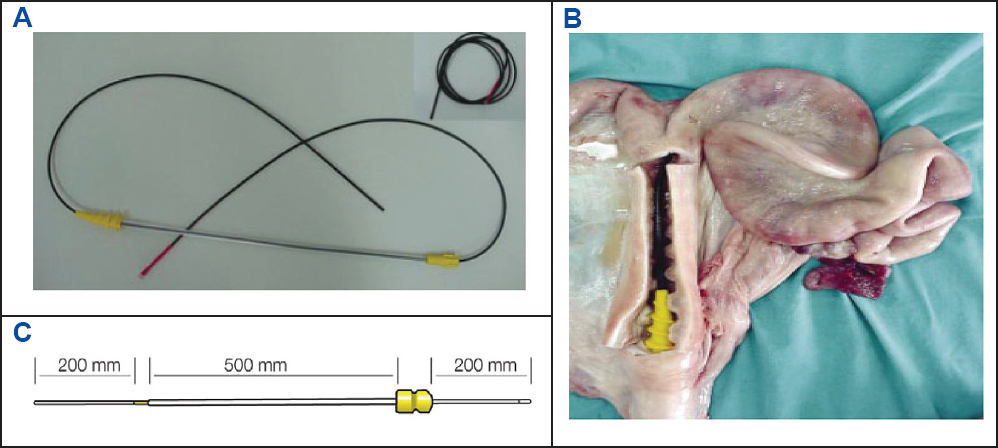
The main obstacle to the trans-cervical AI is the presence of folds in the cervical canal (194). In our laboratory, we use a deep insemination device, called the Deepglodenpig™ catheter (IMV technologies). It involves a standard catheter through which a narrow soft inner tube (4 mm outer diameter) passes and extends 200 mm beyond the tip of the outer catheter and penetrates the cervical canal to lie in the body of the uterus or the posterior horn (Fig. 6-8). The tip of the deep inseminating device carries a smooth rounded white plastic plug with side holes to permit the passage of diluted semen (Fig. 6). On withdrawal, the white tip enables the immediate identification of any blood. The procedures of IUI with liquid-stored semen are shown in Fig. 9. More than 95% of the sows can be easily inseminated with this device and if some resistance is felt during introduction of the catheter, it is necessary to withdraw and reposition of the catheter. On average, the whole insemination procedure takes no more than 5 min, and the placing of the device takes no more than a minute. Although IUI with liquid-stored semen is currently used by practitioners in the EU and USA, sperm numbers per AI dose is not yet standardized. It appears that the threshold dose for achieving a systematic high fertility success is at least 1x109 sperm. However, when farm condition or semen quality is suboptimal, it is prudent to use 1.5x109 spermatozoa per IUI dose (11, 85, 195-198).
Figure 7. The full length of the intrauterine catheter “Deepgoldenpig™” (A) at the stage of insemination in the reproductive tract of the sow (B); adapted from IMV technologies (France) and Watson and Behan (89).
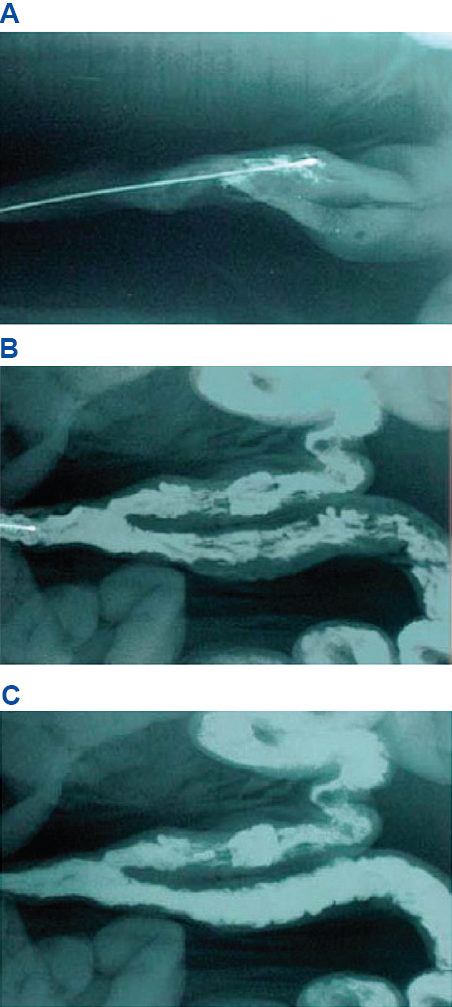
Figure 8. Radiographs of the intrauterine catheter at the beginning of insemination (A) and after 1 (B) and 2 (C) min of insemination; adapted from IMV technologies (France)
Figure 9. Procedures of intrauterine insemination of a sow which include removal of the Deepgoldenpig™ catheter from the plastic cover (A), bringing the semen doses in GTB bags (801 and 402 ml; B), careful insertion of the lubricated catheter in the genital tract (C), direct connection of the semen bag to the catheter (D) and discharging the semen by making a gentle pressure on the bag (E-F); adapted from IMV technologies (France).
DIUI with frozen-thawed semen
To the authors’ knowledge, few field trials have been published for evaluating the potential effectiveness of IUI with cryopreserved sperm, and fertility outcomes are not encouraging (11). Nonetheless, DIUI offers optimistic possibilities to an effective and efficient use of frozen-thawed semen (194, 196, 198, 200). In fixed-time AI protocols, a single DIUI close to the beginning of ovulation with 1 x109 frozen-thawed spermatozoa (~ 500-560x106 live sperm per dose) can achieve high fertility outcomes (farrowing rates over 80% with > 10 piglets born per litter) (11, 201). Moreover, a second fixed-time AI or doubling sperm numbers in single fixed-time AI partially compensates some variations in the expected time of ovulation (11).
Different devices are available for DIUI (Fig. 10) without sedation of the animals (197). After thorough cleaning of the perineal area of the sows, an AI spirette (Minitüb, Germany) is inserted through the vagina into the cervix and used to manipulate a specially designed flexible catheter (working length 1.80 m, outer diameter 4 mm, diameter of the inner tubing 1.8 mm) (Fig. 10). The flexible catheter is then inserted through the spirette, passed through the cervical canal and moved forward along one uterine horn until its total length has been inserted. Before performing DIUI, the inner tubing of the flexible catheter is rinsed with BTS diluent and refilled with ~2 ml of BTS extender at 30°C. The predetermined insemination dose of spermatozoa in 5 ml at 30°C is then infused into one uterine horn using a 5-ml disposable syringe attached to the inner tubing of the flexible catheter. Subsequently, an additional 5 ml of BTS extender is used to force all remaining spermatozoa out of the flexible catheter (197).
Figure 10. A flexible catheter used for deep intrauterine insemination with frozen-thawed boar semen (A), the silhouette of the catheter inserted into a uterine horn (B); adapted from Martinez et al. (197). A commercial device (GT Deep™, IMV technologies) used for uterine horn insemination in sows (C).
The unique characteristics of boar sperm metabolism render AI in pigs more popular compared with other livestock species. The key to the widespread application of AI worldwide is the ability to preserve sperm fertility for up to a week at nearly room temperature, using simple procedures of semen collection, storage and AI. The use of sperm-rich fraction, holding of semen at 18°C and fortification of the storage extender with a combination of milk proteins and egg yolk emulsified in presence of Equex STM and BHT, can protect boar sperm during chilled storage at 5°C; however, further studies are needed to evaluate fertility of chilled semen versus that of stored semen at 16-20°C. In spite of the availability of freezing technology, frozen semen is not widely used in the commercial pig production, mainly because it is not as cost effective as liquid semen. Development of an efficient protocol for synchronizing the time of ovulation in sows and gilts coupled with improving DIUI technique will make a breakthrough in the commercial use of frozen semen. In order to improve fertility of liquid semen obtained from a subfertile terminal sire with good genetics, we suggest the following recommendations: (a) increase the hygienic standards during semen collection to minimize bacterial contamination of the ejaculate, (b) collect the sperm-rich fraction, (c) select the appropriate storage extender and treat the semen with antibiotics based on a bacterial resistance test, (d) select good quality spermatozoa through colloidal purification of semen, (e) add caffeinated media to semen doses just before AI, and (f) carefully deposit semen doses in the uterine body during AI.
© 2014 Khalifa T. This is an open-access article published under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This work was supported by Nidacon International AB, Flöjelbergsgatan 16B, SE-431 37 Mölndal, Sweden (Project # 1003-P; 2011-2012) and by the Veterinary Research Institute, Hellenic Agricultural Organization, 57001 NAGREF Campus, Thermi, P.O. Box 60272, Thessaloniki, Greece.
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 37, Issue 1, Pages 5-34, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.14432/j.macvetrev.2014.09.022, 2014