Mac Vet Rev 2014; 37 (2): 171 - 177
 10.14432/j.macvetrev.2014.09.023
10.14432/j.macvetrev.2014.09.023
Received: 25 June 2014
Received in revised form: 18 August 2014
Accepted: 29 August 2014
Available Online First: 15 September 2014
Published on: 15 October 2014
Keywords: true anoestrus, ovary, GnRH, eCG, dairy cow
The early resumption of estrous cycles following parturition in dairy industry has a great impact in reaching high reproductive efficiency (27). Anestrus is a physiological state during the postpartum period in high-producing dairy cows, with major economic implications since it increases the length of the calving interval (17). In general, a calving interval of 12-13 months is considered economically optimal for dairy cows (25). In order to achieve such a calving interval, a post-partum cow has to resume ovarian activity and conceive within 85 days after calving (24). Santos et al. (30) reported that in high-producing Holstein herds, 6–59 % of the postpartum cows have not resumed cyclicity by Day 60 after calving. Furthermore, cows that have not ovulated in the first 4–8 weeks after calving or not been observed in estrus during the first 60 d postpartum (classified as anovular or anestrual (12), respectively), have a significantly higher risk of being culled than cows that ovulated or have displayed estrus (25).
In the cycling cows, ovarian follicular growth occurs in a regular wavelike pattern. Each wave is comprised of successive phases referred to as recruitment, selection, deviation, dominance, and atresia (20). Trans-rectal ultrasound monitoring of the ovarian follicles reveals that most estrous cycles in cows have two or three follicular waves (11, 33). The follicular waves are first detectable at 4–5 mm follicles approximately on d 0 and d 10 for two-wave and on d 0, 9 and 16 for three-wave inter-ovulatory intervals (11). During each wave, 7-11 small follicles (4 mm in diameter synchronously enter a common growth phase of about 3 days (10). Following that period, a follicular deviation transpired characterized by dissociation in growth rates and the distinction between the future dominant and subordinate follicles (10), resulting in dominant follicle which will attain an ovulatory diameter (10). The latter suppresses the growth of subordinate follicles, thus preventing the emergence of a new follicular wave (2). Each follicular wave is stimulated by a transient increase in the follicle stimulating hormone (FSH) secretion (1) caused by the regression of the dominant follicle of the previous wave (10). The dominant follicle subsequently acquires luteinizing hormone (LH) dependency for its own continued growth and suppresses FSH secretion, therefore starving the subordinate follicles of sufficient FSH support (1, 10). Upon luteal regression and declining of progesterone (P4), follicular maturation associated with increased estradiol synthesis occur, which trigger a LH surge that causes ovulation of a dominant follicle (18).
Subestrus, cystic ovarian disease, static ovaries (true anestrus) and persistent corpus luteum are the four clinical forms of anestrus reported that impediment the reproductive efficiency (36). Among all, static ovaries defined as ovaries with follicles < 8 mm, absence of corpus luteum (CL) and P4 concentration < 0.5 ng/mL between two consecutive ultrasound examinations 7-10 days apart (7, 17), stands as one of the major ovarian dysfunction in the early postpartum period. The physiology underlying their occurrence still remains unclear, although according to Arthur’s 1989 (3), the reasons might be two-fold: either insufficient secretion of gonadotropins for encouraging the folliculogenesis or the inability of the ovaries to respond to gonadotropins. Wiltbank et al. (34) reported that the absence of ovarian follicles larger than 8 mm in diameter may be observed in animals subjected to severe nutritional restriction. The latter is associated with the absence of pulsatile LH release (14). Different treatment methods for static ovaries treatment have shown inconsistent rate of success in the resumption of ovarian cyclicity, imposing further investigations to achieve satisfactory results (28).
The current study was designed to test the hypothesis that single GnRH (gonadotropin releasing hormone) or eCG (equine chorionic gonadotropin) injection will have influence on the ovarian follicular dynamics in cows diagnosed as being in true anestrous in a dose-dependent manner. Therefore, the objectives of the present study were to evaluate the possibility of inducing follicular growth in cows diagnosed with static ovaries using different doses of single GnRH and eCG injection.
Holstein-Friesian cows from three commercial dairy farms in the Republic of Macedonia were involved in the experiment. In Dairy A, cows were housed in a tie-stall on deep straw bedding, whereas in Dairies B and C cows were housed in a free stall with cubicles. On each farm cows were milked twice daily (morning and the afternoon) with average lactation of 6500 kg, 6100 kg and 6400 kg per 305 days, on Dairy A, B and C, respectively. Of the 298 routinely examined cows using trans-rectal ultrasonography, 67 cows were identified as having inactive ovaries (no CL, follicles < 8 mm, and P4 < 0.5 ng/mL). To verify that no follicular growth in aforementioned cows transpired, 7 days later (referred as d 0 = start of the experiment) another ultrasonic examination as well as the blood serum P4 determination was performed. Follicles larger than > 10 mm were recorded in 9 cows and these cows were excluded from further analysis resulting in 58 cows enrolled in the study. Cows were randomly assigned into three groups: GnRH (Fertagyl, Intervet-Holland)=Group 1 (n=23), eCG (Folligon, Intervet-Holland)=Group 2 (n=23) and Control (n=12) cows that were left untreated. Treated cows were further allocated into two subgroups (SB) per treatment according to the applied doses of; for GnRH-treated group, the SB 1.1 (n=8) was treated with 100 µg and SB 1.2 (n=15) with 250 µg GnRH; for the eCG- treated group, the SB 2.1 (n=8) was treated with 750 IU and SB 2.2 (n=15) with 1000 IU eCG. Trans-rectal ultrasonographic monitoring of the follicular dynamics was done on a daily basis, starting from d 0 until ovulation (defined as disappearance of the largest follicle between two consecutive ultrasound examinations) and once on d 7 after ovulation to confirm presence of the corpus luteum in cows which experienced follicular growth. In cows that failed to resume regular folliculogenesis the ultrasound examination was performed simultaneously as the cows which establish cyclicity and continue every third day until 110 DIM. Daily follicular growth rate and treatment respond interval were estimated in all groups using the following definitions: Daily growth rate (GR, mm/day) – the difference between the largest and the smallest diameter of a follicle divided by the number of days; Treatment respond interval (days) - number of days from the first visualization of the follicle until the follicle reached pre-ovulatory size.
Monitoring of the follicular dynamics was done by transrectal ultrasonography of the ovaries using B-mode ultrasound machine Aloka SSD 500, (Tokyo, Japan) equipped with rectal linear 7.5 MHz transducer, connected with a printer, so that images of each ovary could be taken daily. Before insertion of the lubricated transducer, the rectum was emptied, and the ovaries were first manually located. During each examination and using the prints taken during the ultrasound, ovarian maps were drawn to record the size and relative position of the follicles. The diameters of the follicles were obtained from two linear measurements taken at right angles by means of electronic calipers located on the ultrasound device using the images on which the diameters of the follicles were maximal.
Blood samples were collected on d -7, d 0 and d 7 after ovulation from the coccygeal vein into glass tubes and centrifuged (3000 rpm x g 10 minute) within 3 hours after collection. The samples were stored at – 20°C until determination of progesterone by enzyme-immune assay (EIA) was performed. The determination was done at the laboratory of endocrinology at the Faculty of Veterinary Medicine – Skopje (Macedonia), using a commercially available kit (HUMAN, Progesterone ELISA Test - Germany). The intra- and inter-assay coefficients of variation were <10%. The lower detection limit was 0.07 ng ml-1.
Statistical analysis was performed by assessment of the mean (x), standard deviation (SD), and standard error of mean (SEM) values. Standard T-test was used for all analysis to assess differences between the groups. The results are expressed as mean ± SEM.
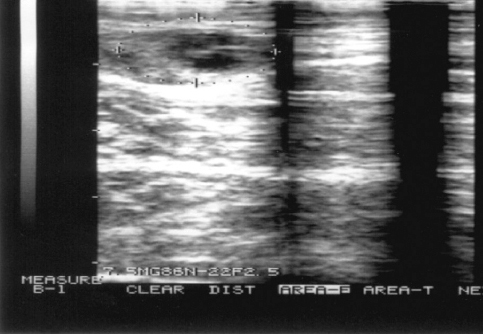
A total of 58 cows (19.46 %) were classified as having static ovaries (Fig. 1) and subjected to further analysis. The average period from calving until the start of the experiment were 72.17 ± 2.22 days, 69.00 ± 1.77 days; 70.62 ± 2.28 days, 73.26 ± 2.7 days and 76.25 ± 3.40 days for SB 1.1, 1.2; 2.1, 2.2 and control group cows respectively. The mean serum concentrations of P4 in cows from all subgroups on d-7 and d 0 were < 0.5 ng/mL (data not shown).
Figure 1. Ultrasound diagnose of static ovary (absent CL, follicles < 8 mm)
Overall, the percentages of cows that established regular follicular growth (defined as continued development of the follicles) in GnRH Group 1, eCG Group 2, and the control group were 56.52 % (13/23), 60.86 % (14/23) and 41.66 % (5/12), whereas 43.47% (10/23), 39.13% (9/23) and 58.33 (7/12) cows failed to respond and to resume follicular growth, respectively. Resumption of the follicular growth occurred in 86.6% (13/15) SB 2.2 cows, 66.6 % (10/15) in SB 1.2, 38.5 % (3/8) in SB 1.1, 12.5 % (1/8) in SB 2.1 cows. The daily follicular growth rate was higher in SB 2.2 (1.6 ± 0.2 mm/day) in comparison to the cows of the remaining SB (1.1, 1.2, 2.1) being (1.26 ± 0.2 mm/day, 1.55 ± 0.4 mm/day, 1.3 mm/day) and the control group being (0.8 ± 0.2 mm/day), respectively. Cows in the SB 2.2 experienced shorter (5.7 ± 0.2 days) treatment respond interval than cows in SB 1.1 1.2, and 2.1 (9 ± 0.57 days, 6.7 ± 0.2 days, and 8 days, respectively) and control group (17 ± 0.7 days). Furthermore, ovulation occurred from the follicles which were larger in control group cows (16.1 ± 0.2 mm) than cows in the SB 1.1 (14.7 ± 0.3 mm), SB 1.2 group (15.6 ± 0.2 mm), SB 2.1 (14.3 mm) and SB 2.2 (15.4 ± 0.1 mm). The diameter of the second largest follicle was higher in SB 2.2 (11.7 ± 0.2 mm) than remaining SB as well as control group (SB 1.1 9.4 ± 0.3 mm, SB 1.2 10.2 ± 0.2 mm, SB 2.1 10.5 mm) and 8.4 ± 0.2 mm, respectively. Cows in SB 2.2 showed a higher rate (42.85 %) of multiple ovulations (defined as disappearance of the co-dominant follicles between two consecutive ultrasound examinations) in comparison with the cows in SB 2.1 (15.38 %) whilst no multiple ovulations were observed in the remaining subgroups.
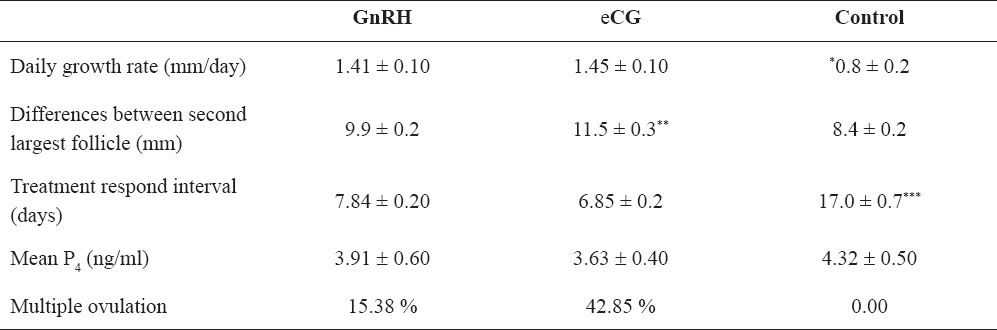
Overall, there were not significant differences in the daily follicular growth rate between cows in both treatment groups (GnRH vs eCG, 1.41 ± 0.1 mm/day vs. 1.45 ± 0.1 mm/day, p > 0.05). However, there was a significant difference compared to the control group (1.41 ± 0.1; 1.45 ± 0.1 mm/day versus 0.8 ± 0.2 mm/day, p<0.001). eCG Group 2 had a numerically faster treatment respond interval than GnRH Group 1 (6.85 ± 0.2 days vs. 7.84 ± 0.2 days), but this was not significant (p>0.05). Moreover, both eCG and GnRH groups had significantly different treatment respond intervals than cows from the control group (17 ± 0.7 days, p<0.001). Significant differences were observed between eCG, GnRH and control groups in the diameter of the second largest follicles being 11.5 ± 0.3 mm, 9.9 ± 0.2 mm and 8.4 ± 0.2 mm, respectively (p<0.05), with no differences between GnRH and controls (p>0.05). The average P4 concentrations on day 7 after ovulation were 3.91 ± 0.6 ng/ml, 3.63 ± 0.4 ng/ml, 4.32 ± 0.5 ng/ml in Group 1, Group 2 and the control group, respectively but no differences were recorded (p>0.05).
Table 1. Difference between GnRH, eCG and control group cows. Asterisks indicate significant differences between groups
The present study compared the effects of single GnRH or eCG injection on the resumption of ovarian cyclicity in true postpartum anestrous cows. In order to initiate treat postpartum anestrous cows for early establishment of cyclic activity and the possibility of conception, the importance of identifying anestrous cows could lead to increased reproductive efficiency in the herd (7). By implementing transrectal ultrasonography accompanied with P4 assay, approximately 20% of the cows were diagnosed as being in true anestrous, which correspondents with the data in the literature 19 % (23); 13% (16).
Treatment with GnRH was followed by the establishment of regular folliculogenesis in 56.52% of the cows. Similarly, when postpartum anestrous cows were treated with GnRH, 50% of those cows resume cyclic activity (19). We suggest that the reduced ovarian response is likely due to lack of LH receptors on granulosa cells (35). Acquisition of the LH receptors has been shown to occur near the selection of the future dominant follicle when the follicle reaches a size of around 8.5 mm (10). Therefore, the presence or absence of the selected dominant follicle at the time of treatment with GnRH apparently determines the success of the resumption of the cyclicity (5). Although systemic administration of GnRH causes increased LH concentration within 2-4 h (32), the latter has been shown to result in reduced response to the subsequent progress of the follicles if this increase occurs before the selection (29). Contrary, if the dominant follicle is already selected, it becomes increasingly responsive to LH and will continue to grow (5). The eCG has both FSH and LH like activity dominantly follicle-stimulating activity, (26); thus parenteral administration stimulates follicular growth and ovulation in cattle (22). Our results have shown that 60.86 % of the treated cows resumed cyclicity after eCG treatment which is in agreement with the results reported by Mansoor et al. (19). In that study 83.3% of the cows establish cyclicity when treated with eCG similarly as subgroup 2.2 (86.6%). The increased response in the current study was achieved using smaller doses of 1000 IU than the dose used in the study of Mansoor et al. (3000 IU eCG) (19). When 1500 IU eCG was used, resumption of follicular growth occurred in 83.3 % of the treated cows (9) which also correspond with the results observed in the present study.
Furthermore, our results have shown that administration of 250 µg GnRH or 1000 IU eCG causes increased ovarian response when compared to lower applied doses (100 µg GnRH and 750 IU eCG). This could lead to assumption that smaller doses of GnRH and eCG may have partially effect of the initiation of the follicular growth in cows with inactive ovaries. However, it should be noted that, in the early postpartum period, most of the cows are unable to meet their energy requirements as a result of decreased feed intake at a time when energy requirements are increasing (31). Therefore, an early pospartum cows enter a period of negative energy balance (NEB) which usually demises after 60 days postpartum (8). Cows in NEB have reduced genes expression in granulosa cell for LH receptors as well as having suppressed pulsatile LH secretion. This in turn causes a reduced ovarian response to LH stimulation (for reviews see Diskin et al.) (6). Therefore, it can be assumed that cows (especially in the subgroups 1.1 and 2.1) might still be in negative energy balance during the experiment and hence had a decreased treatment response.
The daily follicular growth rate did not differ between GnRH and eCG, (1.41 ± 0.1 mm/day vs. 1.45 ± 0.1 mm/day), respectively. However, cows from both treatment groups significantly differed from the cows in the control group (0.8 ± 0.2 mm/day). At the moment there is a lack of data in the literature, although these results could support the hypothesis of insufficient secretion of gonatrophins (3), necessary for enhanced resumption of the ovarian activity. In the current study eCG treated cows showed numerically faster treatment respond interval in comparison to GnRH (6.85 ± 0.2 days vs. 7.84 ± 0.5 days respectively) and both GnRH and eCG treated cows significantly differed from the control group (17 ± 0.7 days). The faster eCG response in comparison to GnRH could be a consequence of the different mode of action of these hormones. Imperfection of the specific receptors for GnRH on the ovaries, demonstrate that its primary effect is on pituitary, stimulating pulsate release of FSH and LH in peripheral circulation (4). On the contrary, dominantly follicle-stimulating activity of eCG directly affects follicles and causes stimulation of folliculogenesis (26).
Due to the increased representation of multiple ovulations (42.85%) in the eCG treated cows, despite having higher follicular growth rate and faster treatment respond interval, the treatment with eCG, cannot be suggested as method of choice for treatment of static ovaries in dairy cows. However, this treatment can be recommended in the beef cows where high rate of twinning is more desirable (15). Similar rates, (47% multiple ovulations) were observed by Musssard et al. (19) pointing that the reason might be result of follicular development during the presence of low progesterone concentration in the absence of CL. It has been shown that CL absent cows has increased plasma estradiol concentration compared to CL present cows (13). The latter could increase LH frequency which probably is involved in the growth of the second largest follicle leading to co-dominance (13). On the other hand, dominantly follicle-stimulating activity as well as the long half-life of eCG (5-7 days) (27), may lengthen the common growth phase of recruited follicles thereby disrupting the deviation process and subsequently allowing development of multiple dominant follicles.
The results of this study revealed several conclusions. First, ultrasound examination in cows more than 60 days after calving, accompanied by serum P4 determination is a reliable method for detecting the static ovaries. Second, the single application of eCG or GnRH enhanced the folliculogenesis through the processes of emergence, selection and dominance and re-establishment of cyclic activity, although the treatment success depends primarily on the energy status of the cow. Third, the eCG response is faster and more successful compared to GnRH, however, due to the increased occurrence of multiple ovulation and the possibility of higher twinning rate, this treatment method should be used without AI. Therefore as a general conclusion, single application of 250 μg GnRH is an acceptable treatment method for static ovaries in terms of encouraging and reestablishing normal follicular growth concomitantly with the possibility of subsequent artificial insemination.
© 2014 Atanasov B. This is an open-access article published under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 37, Issue 2, Pages 171-177, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.14432/j.macvetrev.2014.09.023, 2014