Mac Vet Rev 2015; 38 (2): 167 - 173
 10.14432/j.macvetrev.2015.05.043
10.14432/j.macvetrev.2015.05.043
Received: 28 February 2015
Received in revised form: 20 April 2015
Accepted: 24 April 2015
Available Online First: 04 May 2015
Published on: 15 October 2015
Keywords: dog, dermal papillae, dermal papillae-like tissue, adipose-derived mesenchymal stem cells, hair regeneration
Alopecia is a common dermatological disease in veterinary and human medicine. It may be caused by inflammation, infection, traumatic damage as a result of chemotherapy or radiotherapy, nutritional deficiencies, hormonal or autoimmune disease, or stress. Even though the classification of alopecia in humans is more complex, certain types of human alopecia can be found in canine hair loss (1-3).
The condition, especially in humans, may be mentally stressful to patients going through it due to its effect on their appearance, which is why many different treatment methods have been tried. Out of these, auto hair transplantation has been the most effective. In addition, because of critical limitations due to cost, amount of hair and number of operations (4), studies on hair regeneration have been attempted using cultured hair follicle cells or mesenchymal stem cells (4-8).
Hair growth is a result of active epidermal and dermal interactions (6, 9-14). In general, the dermal component is thought of as an inducer and the epidermal component as a responder in the process of hair growth (15). Dermal papilla (DP) is a cluster of highly specialized fibroblasts located at the base of the hair follicle (HF). It is believed that DP not only determines the length, thickness and shape of the hair shaft (16), but also regulates hair follicle development and growth. For this reason, active research on DPs has been performed.
Hair regeneration has been attempted using tissue and cellular recombination of the epidermal and dermal component (15, 17). However, DP isolation is technically laborious and difficult, and cultured dermal papilla cells (DPCs) lose their hair inductive abilities after a few passages. To overcome these limitations, studies on the reconstruction of dermal papilla-like tissues have been performed with human umbilical cord-derived mesenchymal stem cells (4, 5, 7).
This study represents the first reconstruction of DPLTs using canine adipose-derived mesenchymal stem cells (AD-MSCs); we compared them to actual canine DPCs in order to evaluate DPLTs as an alternative to actual DPs. We expect the findings of this study to broaden our understanding of the morphological and molecular characteristics of canine DPCs, and to represent a valuable source for the investigation of canine and human hair regeneration.
We used canine AD-MSCs that were supplied from Seung Hoon Lee (Department of Veterinary Clinical Sciences, College of Veterinary Medicine, Seoul National University); these AD-MSCs were previously characterized (18). The AD-MSCs were cultured in low-glucose Dulbecco’s modified eagle medium (DMEM, Gibco®, Life Technologies, Grand Island, NY, USA) with 10% fetal bovine serum (FBS, Gibco®, Life Technologies, Grand Island, NY, USA), 100 units/mL penicillin and 100 mg/mL streptomycin at 37.0 °C, in a 5% CO2 incubator.
Full-thickness skin biopsies obtained from the flank region of healthy beagle dogs approximately 3 years old were immediately placed in phosphate-buffered saline (PBS). Under a dissecting microscope, the hair follicles were isolated using a No. 15 blade, watchmaker’s forceps, and 23G needles. Isolated hair follicles were then transferred to a fresh medium-containing dish for further isolation. After the hair cycle stage was evaluated by the position of DP and tip of hair shaft, DPs in late anagen were isolated (19). Five DPs were placed together in a single well of a 12-well cell-culture cluster, which contained 1.2 mL of AmnioMAX™-C100 complete medium (AmnioMAX™, Invitrogen, Carlsbad, CA, USA) supplemented with 10% inactivated and steroid-free canine serum at 37 °C, in a 5% CO2 incubator (6, 8, 20-22). The culture medium was replaced every third day. Primary cultured DPCs were harvested when confluent and then passaged on every tenth day. All procedures were approved by the Seoul National University’s Institutional Animal Care and Use Committee (SNU-140729-5).
Canine AD-MSCs were subjected to a monolayer culture in DMEM supplemented with 10% FBS, 100 units/mL penicillin and 100 mg/mL streptomycin, until the cells occupied approximately 80% of the culture dish. Next, the culture medium was replaced with dermal papilla-forming medium (DPFM) (23) that contained 10 ng/mL hydrocortisone, 5 mL Insulin-Transferrin-Selenium liquid media (ITS, Gibco®, Life Tecnologies, Grand Island, NY, USA), and 100 units/mL penicillin, 100 mg/mL streptomycin, and 20 ng/mL HGF (recombinant human HGF, Gibco®, Life Tecnologies, NY, USA). The medium was changed every third day for three weeks, after which the culture was treated with the StemPro® Accutase® Cell Dissociation Reagent (Accutase®, Gibco®, Life Technologies, Grand Island, NY, USA) at concentrations ranging from 20 to 40 μL/cm2 to detach the cells from the culture dish (detach-attach step). Cell distribution was changed and cell aggregation was observed 24 h after treatment with Accutase; the aggregated cell mass became suspended in the medium five to seven days after the treatment. The suspended cell aggregates were then isolated from the culture by centrifugation at 500 rpm for 3 min. All procedures were performed based on those of previous studies (4, 23).
Total RNA from all three cell types (canine AD-MSCs, DPCs and DPLTs) was isolated using Hybrid-R™ with RiboEx® (GeneAll®, Seoul, Korea) according to the manufacturer’s instructions. cDNA was then synthesized, using one microgram of the total RNA, with a PrimeScript™ RT-PCR Kit (Takara Bio Inc., Shiga, Japan) for RT-PCR processing according to the manufacturer’s instructions. Cycling conditions included initial denaturation for 10 min at 94 °C, followed by thirty-five 15-s cycles at 95 °C and 60-s cycles at 60 °C. The products were then analyzed by electrophoresis on a 1.6% agarose gel and subsequently visualized by the Redsafe™ Nucleic Acid Staining Solution (iNtRON Biotechnology, Gyeonggi-do, Korea). We tested well-known hair-inductive markers of canine DPCs, such as alkaline phosphatase detection (ALP), Wnt inhibitory factor 1 (WIF1), laminin, vimentin, versican, and prominin-1 (1, 8, 13, 15, 24). Glyceraldehyde 3-phosphate de hydrogenase was used as the reference gene.
The cell and tissue extracts were prepared using the PRO-PREP™ protein extraction solution (iNtRON Biotechnology, Gyeonggi-do, Korea). Protein samples were run on a 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel, transferred onto a nitrocellulose membrane, and incubated with an anti-human versican (VCAN) antibody (R&D systems, Minneapolis, MN, USA), anti-laminin antibody (ab11575, Abcam, Cambridge, MA, USA), and anti-vimentin antibody (ab8069, Abcam, Cambridge, MA, USA). An anti-beta actin antibody was used (sc-47778, Santa Cruz Biotechnologiy, Paso Robles, CA, USA) as an internal control. Blots were then incubated with a peroxidase-conjugated secondary antibody and visualized using a Novex® ECL Chemiluminescent Substrate Reagent Kit (ECL, Novex®, Grand Island, Life Technologies, NY, USA).
A student’s t-test was used for comparing the size between the canine DPCs and DPLTs. A value of p<0.05 was considered to be statistically significant. The statistical analyses were performed using GraphPad Prism version 6.00 for Windows (GraphPad Software, La Jolla, CA, USA).
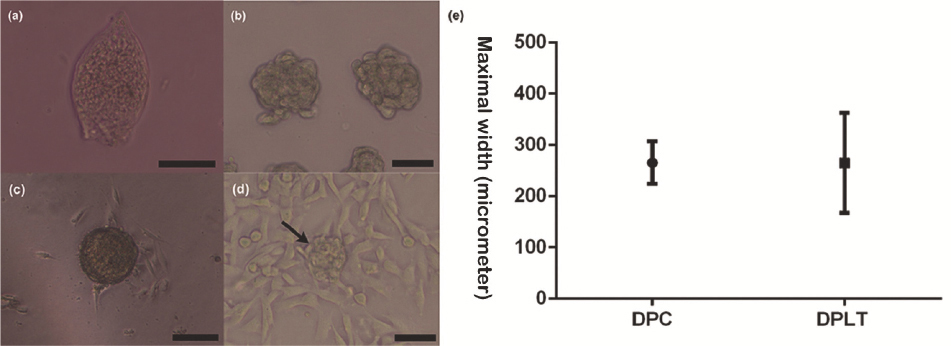
The DPs were teardrop-shaped (Fig. 1a). Their approximate sizes were measured and compared with those of DPLTs (Fig. 1b). The mean actual canine DP maximal width was 265.8 ± 11.50 μm. The first outgrowth of dermal papilla cells presented after five days, and the cells had a flattened, polygonal morphology (Fig. 1c) with multiple cytoplasmic processes. We also observed the aggregative growth pattern in the multi-layer culture with the formation of the pseudo-papillae (Fig. 1d) and cells formed multilayered conglomerates on day 8.
Figure 1 Dermal papilla (DP), cultured dermal papilla cells (DPCs) and dermal papilla-like tissues (DPLTs) (a). Isolated DP (200X) (b). Reconstructed DPLTs from canine AD-MSCs (100X) (c). Primary DPCs culture. The first outgrowth cell was seen on day 5 (100X) (d). Typical multilayered, aggregative DPC growth pattern and the formation of pseudopapaillae (arrow) (100X) (e). Comparison of DPs and DPLTs: There was no statistically significant difference in the maximal width between two groups
After the cell detach-attach process, cell aggregates were formed within five days. The mean DPLT size was 296 ± 12.43 μm, and there was no statistical difference when compared to actual DP size (p = 0.1465) (Fig. 1e). Various factors affected DPLT formation; of these, cell inoculation density was one of the most prominent factors. We tried DPLT formation with different cell inoculation densities, from 2.0 x 104 cells/cm2 to 8.0 x 104 cells/cm2. It was found that the optimal inoculation cell density was from 6.0 x 104 cells/cm2 to 8.0 x 104 cells/cm2 (data not shown). Cell migration only occurred in densities lower than 2.0 x 104 cells/cm2. Even though significant aggregation was observed from a density of 4.0 x 104 cells/cm2, in the optimal density, the number of DPLTs was higher and the sizes were more regular than those of DPLTs in a density of 4.0 x 104 cells/cm2.
We performed RT-PCR and Western blotting to evaluate molecular similarities between DPLTs and DPs. Well-known hair-inductive markers, as previously mentioned, were assessed for evaluating the hair-inductive abilities of three groups on a genetic level. In addition, we assessed the quantitative protein expression level with laminin, vimentin and versican.
According to previous studies, ALP was used as an indicator for hair inductivity (6, 15), and versican was expressed in anagen hair follicles but absent in telogen hair follicles. This might support the idea that DP played an important role in anagen induction and maintenance (15, 24). WIF1, vimentin and laminin were related to signal transduction, cellular junction and cytoskeleton, and extracellular matrix and cell adhesion of DPs, respectively (12). In previous studies, WIF1 was considered to positively relate to in vivo murine hair-inductive capacity and was significantly over-represented in freshly isolated canine DPs (1, 6, 11, 13). Vimentin is known for the functions of promoting the proliferation of DPCs and increasing cell migration and growth factor expression and laminin plays an important role in promoting DP development and function during early hair morphogenesis (25).
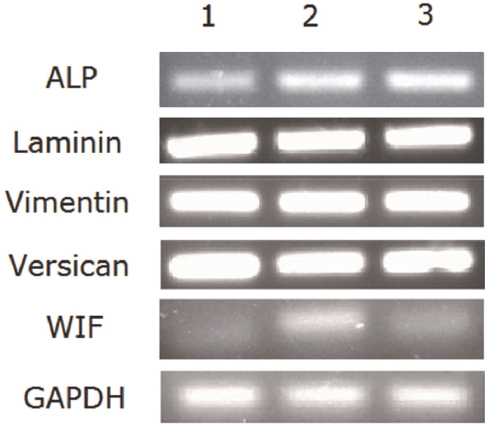
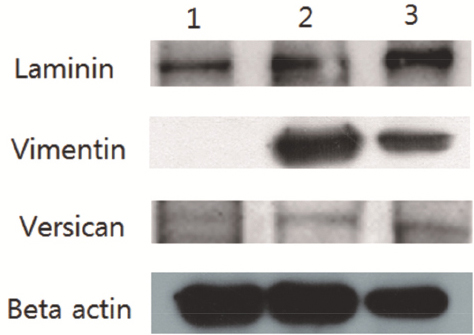
RT-PCR analysis revealed that AD-MSCs, DPCs, and DPLTs expressed all markers as mentioned above (Fig. 2). In western blotting, all three groups showed similar versican expression level, which was reflected by their hair-inducing ability. However, we also observed that vimentin and laminin had a higher level of expression in DPCs and DPLTs compared to AD-MSCs (Fig. 3), which might mean that DPLTs have higher hair inductivity than AD-MSCs and the potential to be a viable substitute for actual DPs.
Figure 2 RT-PCR of reconstructed DPLTs. Lane 1, monolayer cultured canine AD-MSCs (third passage). Lane 2, third passage of cultured canine DPCs. Lane 3, DPLTs reconstructed from the third-passage canine AD-MSCs. Hair-inductive markers, ALP, WIF1, laminin, vimentin, versican, and prominin-1, were similarly expressed in all three groups
Figure 3 Western blot of reconstructed DPLTs. Lane 1, monolayer cultured canine AD-MSCs (third passage). Lane 2, third passage of cultured canine DPCs. Lane 3, DPLTs reconstructed from third passage canine AD-MSCs. Vimentin and laminin had a higher level of expression in DPC and DPLT than in AD-MSCs
Limited research has been done on canine hair follicles, and our understanding of hair follicles in veterinary and human medicine relies heavily on murine studies (14, 15, 26). Not only are canine primary anagen hair follicles morphologically analogous to human anagen hair follicles in terms of size and structure but dogs also develop most major human alopecia (2, 27). Therefore, canine hair follicles could be a helpful model for the investigation of human hair follicle biology.
There have been many trials and successes of regeneration of hair with epidermal and dermal components, both in vitro and in vivo (15). Yoo et al. (23) also observed hair regeneration on the scalps of mice through transplantation of epidermal and mesenchymal compartments. The difference was that they applied aggregates from human umbilical cord-derived mesenchymal stem cells as an alternative to the dermal components comparing to former studies.
DP plays an important role not only in regulating hair follicle development and growth, but also as a reservoir of mesenchymal stem cells. Driskell et al. (27) noted that DPC expansion and generation might be one way of strategically treating alopecia. However, cultured DPCs lose their thrichogenic properties in vitro after few passages (15, 27, 28). Reconstructing DPLTs is one of the ways to overcome that limitation.
In this study, we found that the expression level of hair-inductive markers and proteins between DPCs and DPLTs showed similar patterns, which suggested that cell aggregates reconstructed from AD-MSCs might be excellent substitutes for actual DPs. DPLTs may have the potential to be a key structure in a future treatment method of alopecia that does not respond to medical treatments, and in auto-transplantations. We may also consider a new trial to treat alopecia in veterinary medicine.
In previous study, it was demonstrated that AD-MSC, which had various cytokine-secreting properties and paracrine functions, showed an effect on hair growth by promoting the survival and proliferation of DPCs (29). In addition, Osada et al. (30) demonstrated that aggregates made from later passage DPCs (more than 10 passages) did induce new hair follicles unlike dissociated DPCs that lost their hair follicle-inducing ability at later passage (over passage 4). Considering the results of previous and present studies, we cautiously speculate that DPLTs would have a direct effect on hair re-growth as an alternative structure of DPs and show analogous behavior patterns in vivo or under epidermal-mesenchymal interactions (EMIs) compared to dissociated AD-MSCs.
In the culture of canine DPCs, they showed a typical growth pattern and morphology, as in previous studies (1, 21, 30). We hypothesized that early-passage canine DPCs cultured in AminioMAX had the same capacity for hair induction as canine DPs, based on previous studies (1, 21). We used third-passage DPCs and observed their aggregative culture patterns. In the DP microdissection procedures, we selected late-anagen-phase hair follicles based on morphological criteria; to make the procedures simple, we chose large hair follicles, which could explain why DP sizes were measured larger than expected.
In conclusion, in this study, we reconstructed canine DPLTs with AD-MSCs, which are a more easily accessible source than other mesenchymal stem cells and DPCs, and evaluated them as alternatives to canine DPs. We believe that DPLTs may form the basis of potential therapeutic applications. However, further investigations on DPLTs under active EMIs or in vivo should be performed.
Acknowledgment
This research was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF), funded by the Ministry of Education, Science and Technology (2013-011357).
Copyright
© 2015 Bae S. This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial License (http://creativecommons.org), which permits unrestricted non-commercial use, distribution, and reproduction in any medium provided the original work is properly cited.
Conflict of Interest Statement
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Citation Information
Macedonian Veterinary Review. Volume 38, Issue 2, Pages 167-173, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.14432/j.macvetrev.2015.05.043, 2015