Mac Vet Rev 2015; 38 (2): 189 - 193
 10.14432/j.macvetrev.2015.07.049
10.14432/j.macvetrev.2015.07.049
Received: 30 April 2015
Received in revised form: 10 June 2015
Accepted: 16 June 2015
Available Online First: 06 July 2015
Published on: 15 October 2015
Keywords: Holstein cows, biochemical parameters, transition period
Production diseases are metabolic disorders in animals with genetically determined high production capacities, based on disbalance between the animal’s input (feed utilization) and output (animal’s product, e.g. milk). In dairy cows this disbalance is evident usually during the so-called “transition period” (3 weeks prior and 3 weeks after calving) (1), when very significant changes in the hormonal status occur, which leads to redirection of metabolic pathways, i.e. favoring of anabolic processes in the mammary gland (milk compounds production) vs. metabolic processes in other tissues. These high production performances also cause high nutrition demands, which exceed the animal’s metabolic capacities (inadequate feed utilization), which usually leads to an aberrant physiological state known as negative energy balance (NEB) (2). Developing of NEB is closely related with specificities of the ruminant’s metabolism. In ruminants, alimentary glucose is converted by ruminal microflora into short-chain volatile fatty acids, and only some of them are substrate for gluconeogenesis which occurs in liver, and is the main source of serum glucose in ruminants. This causes blood glucose in ruminants to have a relatively lower level compared with non-ruminants, and it ranges from 2.2 to 3.3 mmol/L (3). There are physiological variations in the glucose level in different phases of the production-reproduction cycle. In normal cows, at the end of the gestation the glucose level is within its physiological range for maintaining of the gluconeogenesis process (4). During the calving the glucose level rapidly increases, which is explained as a result of stress and hormonal status changes (5, 6). After calving, during the early lactation, the glucose level is significantly lower compared with the period before calving (7, 8), and this could be considered as introduction to NEB where synthesis of milk compounds in the mammary gland exceeds the gluconeogenesis capacities of liver, and leads to development of metabolic (production) diseases (8, 9). Increased utilization of glucose by the mammary gland vs. decreased glucoplastic compounds input and decreased gluconeogenesis cause development of hypoglycemia, which very often could be associated with a disorder of the lipid metabolism (7, 10). In cows with hepatic lipidosis gluconeogenesis is decreased, hepatic glycogen pools are depleted, which causes hypoglycemia (11, 12). All these changes in the glucose and lipid metabolisms are also related with hepatocyte morpho-functional integrity.
Animals typically response to NEB by mobilizing body fat reserves, as an attempt to maintain energy homeostasis, so non-esterified fatty acids (NEFAs) are mobilized from the adipose tissue and utilized by the liver, where they could be fully oxidized to CO2, converted to ketone bodies, mainly in beta-hydroxybutyrate (BHBA), or re-esterified into triglycerides. Further, these triglycerides are delivered into the blood as very low density lipoproteins (VLDL), or stored as cytosolic lipid droplets (1). Like other ruminants, dairy cows’ liver has very low capacities of VLDL synthesis and secretion (13), which makes these animals very susceptible to lipid metabolism disorders, manifested as hepatic lipidosis or fatty liver (retention of high levels of triglycerides in hepatocytes), or as ketosis (high concentration of ketone bodies in the blood) (14).
It could be summarized that metabolic disbalance which occurs during the transitional period in dairy cows is a real, but often encrypted problem, with significant implications for milk production, causing enormous economic losson one side, and on the other it can be a predisposition for many clinically manifested health problems, like milk fever, ketosis, retained fetal membranes, metritis (1), abomasum displacement (1, 15) and mastitis (16, 17). In looking at the multifactorial regulatory mechanisms in energy turnover in high-yielding dairy cows during the transition period, the aim of this survey was a monitoring of the metabolic profile during the transition period and detection of the intensity of morphological injury of the hepatocytes.
The examination included 10 (n=10) Holstein cows, that were 3-6 years of age, with an average body weight 450-650 kg. All cows were kept at free stall system, managed and fed under the same conditions.
Blood samples were taken by puncturing of the jugular vein, -5, 5, 10, 30 and 60 days relative to calving. Blood samples were taken into sterile serological tubes and after the spontaneous coagulation, the serum was separated with Sanyo Mistral 2500 centrifuge on 2500 rpm. The serum samples were stored on -18°C ДO -20°C till the analysis. Analyzed biochemical parameters included parameters that are related to carbohydrate and lipid metabolism (glucose, triglycerides, total cholesterol, non-esterified fatty acids - NEFA, and β-hydroxybutirate - BHBA), as well as parameters related to liver functional status (albumin, total protein, urea and total bilirubin).
The level of glucose was determined in the whole blood, immediately after sampling with commercial strips, using Accu-Chek (Roche, USA) glucometer.
Triglycerides, total cholesterol, albumin, total protein, urea and total bilirubin were determined by commercial kits, Human (Germany). NEFA and BHBA were analyzed by Randox (UK) commercial kits. According to the manufacturer’s instructions, the methods were “end-point” and kinetic-enzymatic reaction on photometer STAT FAX 3300 (Awareness Technology Inc, USA).
Body Condition Score was estimated according to Elanco Animal Health Bulletin AI 8478, at days -10, 10, 30 and 60 relative to calving.
Milk production was recorded by an automatic milking system, DeLaval, Sweden at days 10, 30, and 60 relative to calving.
Biochemical parameters presented in Tables 1 to 4, include the dynamics of biochemical parameters starting from day 5 before to day 60 after calving. Results for BCS and milk production are presented in Table 5.
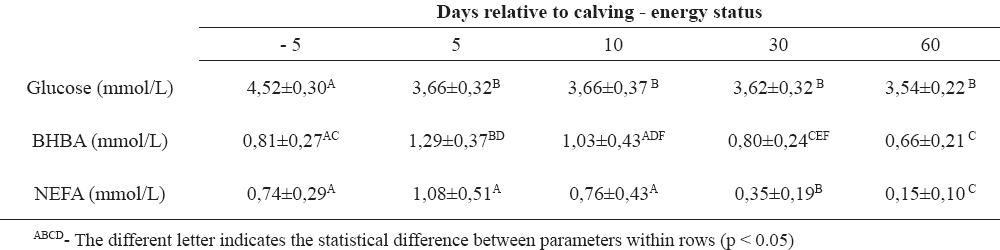
Table 1. Biochemical parameters (X̅±SD) related to energy status in a Holstein dairy cows (n=10)
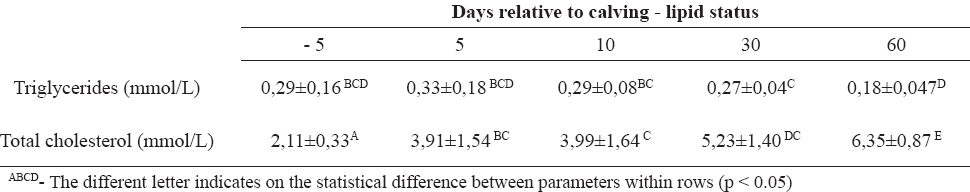
Table 2. Biochemical parameters (X̅±SD) related to the lipid status in Holstein dairy cows (n=10)
Table 3. Biochemical parameters (X̅±SD) related to the hepatic status in Holstein dairy cows (n=10)
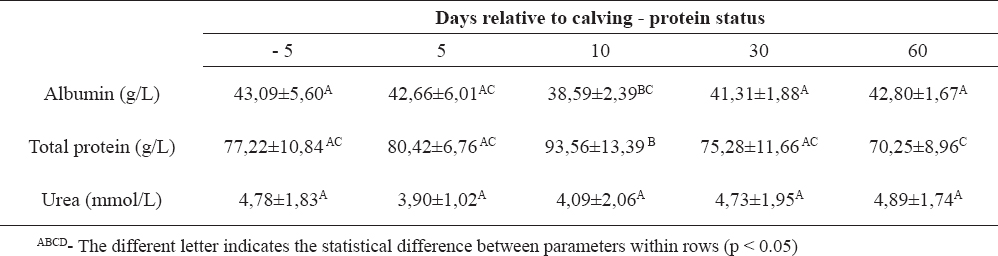
Table 4. Biochemical parameters related to the protein status in Holstein dairy cows (n=10)
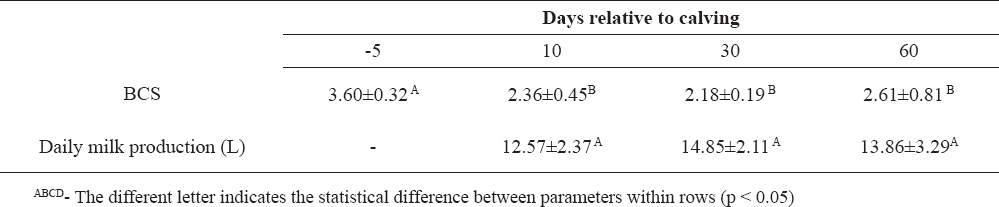
Table 5. BCS and milk production in Holstein dairy cows (n=10)
Dairy cows with high genetic merit for milk production, during the peripartal period change the serum glucose level in different stages of the reproduction cycles (1, 11). We have studied the dynamics of biochemical parameters from day 5 before to day 60 after calving, in order to establish if a cow is capable to maintain those parameters within the physiological range, despite the challenges she is exposed to. Glucose serum concentrations depend on many factors, especially neuro-hormonal regulation and proper feeding management (3). Normoglycemia was observed in the examined cows during the antepartal period, indicating supply of energy precursors from alimentary recourses. Onset of lactation caused hypoglycemic condition due to enhanced utilization of glucose in galactogenesis. There was significant decrease of glucose serum concentrations combined with increased milk production. During the early lactation, the energy balance was switched in udder synthetic processes. NEFA are a “direct indicator” for lipomobilisation (18) and a very dynamic parameter. Subcutaneous and visceral fatty tissue broke down, releasing NEFA which are used in peripheral tissues as an energy source, or they become ingredients of the milk fat (4, 7). Besides the normal glucose status before calving, higher, compared to physiological, serum concentration of NEFA during the antepartal period revealed early lipolysis, as a result of adaptation to high energy demand for fetus growth and milk producing. (5, 9) Early stage lactation is characterized by intensive lipomobilisation for udder anabolism. (6, 14) Significant decrease of NEFA during 30 and 60 days postpartum means balanced metabolic requirements, form alimentary recourses and milk producing. Another parameter for energy status is BHBA, as an “indirect indicator” for negative energy balance. (17, 18) Obtained results for serum BHBA concentration during the antepartal period revealed intensive ketogenesis in hepatocytes, due to inappropriate oxidation of non-esterified fatty acids. BHBA follow the trend of NEFA serum concentration, so the highest values of BHBA were at days 5 and 10 after parturition, but on days 30 and 60 values return in referent ranges. Triglyceridemia indicates the capability of liver for synthesis apolipoprotein B-100, VLDL for elimination of triglycerides in the blood stream during the whole peripartal period, but diminished concentration was significant on day 60 after calving, when NEFA concentrations were very low. These results agree with some previously published results from other investigators (8, 10). Hypocholesterinemia was maintained during the antepartal period, as a result of low “de novo” synthesis of cholesterol. During the postpartal period, serum cholesterol concentration significantly increased, thus 30 and 60 days after calving they reached the highest value. Also, the liver ability for synthesis of apolipoproteins for cholesterol transport was maintained and the highest significant cholesterol concentration was reached 30 and 60 days after calving. Total bilirubin as relevant indicator of liver impairment, (6) during the peripartal period showed an increased value at day 5 relative to calving, because the pathobiochemical processes of gluconeogenesis, glucogenolysis, phosphorilation, beta oxidation happened simultaneously in hepatocytes. Protein status through albumin and total protein concentrations showed changes at day 10 after calving. There were significant increases of protein concentrations at day 10 after calving, with a significant decrease of the level of albumin at the same time period, probably due to decreased synthesis in the liver and increased utilization in the udder.
Biochemical parameters can be used as reliable indicators of degree and duration of metabolic distress in Holstein cows. During the transition period, the lowest value of BCS is achieved at day 30 after calving due to prolonged lipolysis in those cows. Cows included in our study did not achieve their genetic merit for milk production. Although the examined cows were clinically healthy, changes in some biochemical parameters indicate liver increased activity and metabolic stress that could lead to decreased milk production, impaired reproductive performance and, finally, to illness.
Copyright
© 2015 Celeska I. This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial License (http://creativecommons.org), which permits unrestricted non-commercial use, distribution, and reproduction in any medium provided the original work is properly cited.
Conflict of Interest Statement
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Citation Information
Macedonian Veterinary Review. Volume 38, Issue 2, Pages 189-193, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.14432/j.macvetrev.2015.07.049, 2015