Mac Vet Rev 2015; 38 (2): 209 - 216
 10.14432/j.macvetrev.2015.08.052
10.14432/j.macvetrev.2015.08.052
Received: 15 June 2015
Received in revised form: 28 July 2015
Accepted: 07 August 2015
Available Online First: 13 August 2015
Published on: 15 October 2015
Keywords: traditional cheese, enterococci, enterocins, antimicrobial activity
The presence of pathogens in the dairy industry posses a potential risk and a constant concern in the field of food safety for consumers, food business operators and government authorities. A number of food borne disease outbreaks have been associated with these products (1). Although most of these outbreaks were closely related with the consumption of dairy products made from raw milk, post-processing contamination must be taken into account as an important risk factor in the manufacture of such products (2).
Enterococci as a part of lactic acid bacteria (LAB) present a complex, divergent and significant group of bacteria in terms of their interaction with food and humans. Enterococci are omnipresent bacteria and are dominant residents of the digestive tract of humans and animals but their presence it’s not unusual in the surrounding environment like soil, surface waters, plants and vegetables. They can also be found in food, especially in cheese (3). They are found in a high percentage as a part of the dairy microflora. They are especially found in a high percentage in many artisanal made cheeses traditionally produced in the countries from the Mediterranean region, mostly from raw ewe’s or goat’s milk. There is a widely spread opinion that these types of bacteria initially became contaminants from animal organic waste, water or milking equipment and storage tanks, and consequently became an essential component of artisanal cultures (4). These types of bacteria have a fundamental role in the ripening process of traditionally made cheeses (5). As a contribution to their involvement in the unique and specific peculiar taste and flavor, the enterococci also have the ability to protect from various pathogens (6). Recently, the scientific community has increased its interest in enterocins, encouraged by the fact that they act against food-borne pathogens, mostly against L. monocytogenes, S.aureus, B. cereus and vegetative cells and spores of C. botulinum (7, 8).
Lactic acid bacteria for centuries have been used in food and feed conservation and their preservative effects are mainly due to the decreasing of pH values and the formation of organic acids, principally lactic acid (9, 10, 11). Bacteriocins are ribosomally synthesized polypeptides that possess antimicrobial activity and are commonly pH and heat tolerant and are rapidly digested by proteases in the human digestive tract due to their proteinaceous nature (12). Bacteriocins can also inhibit adulteration of food by preventing the outgrowth of non-pathogen and pathogen bacteria. The bacteriocin or bacteriocin-producing LAB except cytolysin, can be used as safe alternatives to chemical preservatives in foods (11, 13), because they are harmless to eukaryotic cells.
Beside the clear evidence that bacteriocinogenic enterococci and their bacteriocins are well known in the literature, the exploration in this field is still an actual subject of the research, as the isolation and characterization of the enterocin producing enterococci from different geographical areas, which have tradition for making artisanal dairy products, could perhaps give new data about the diversity and ecology of Enterococcus strains and their enterocins.
The objectives of this study were to identify bacteriocinogenic enteroccoci isolated from Macedonian traditional cheeses, as well as to determine the existence of bacteriocin structural genes.
A collection of 13 Enterococcus strains that were previously isolated from traditional white pickled (brined) cheese from different regions of Macedonia were tested for presence of bacteriocin structural genes. The enterococci were isolated on a selective agar (KAA, Oxoid, UK) and identified by polymerase chain reaction (PCR) amplification of a part of 16S rRNA gene (14). Species determination was performed using PCR protocol for species-specific enterococcal targets (14, 15). Additionally, isolates negative on species-specific PCR were further determined by BLAST analysis of the 16S rRNA sequences, using available sequence data from GenBank.
The following cultures were tested for sensitivity to enterocins: L. monocytogenes (NCTC 11994), L. innocua (cheese isolate), S. aureus (cheese isolate), P. aeruginosa (NCTC 10662), B. cereus (NCTC 7464), S. enteritidis (meat isolate), E. coli (NCTC 9001) and Y. enterocolitica(ATCC 11303).
For the detection of antimicrobial activity, 50 μl of an overnight culture of the indicator strain was added to 5ml molten soft semisolid Plate count agar (PCA), mixed and poured onto a solid PCA agar plate. A single colony of each enterococcus to be tested for antimicrobial activity was transferred with a sterile loop. Agar plates were incubated for 24 h at 37°C in aerobic condition. The antimicrobial activity was visually detected by observing clear zones around the tested strain. To test the proteinaceous nature of the inhibitors, 10 μl of trypsin solution was deposited on the side of each spot of bacterial growth. The absence of inhibition in the trypsin-spotted zone indicated protease sensitivity.
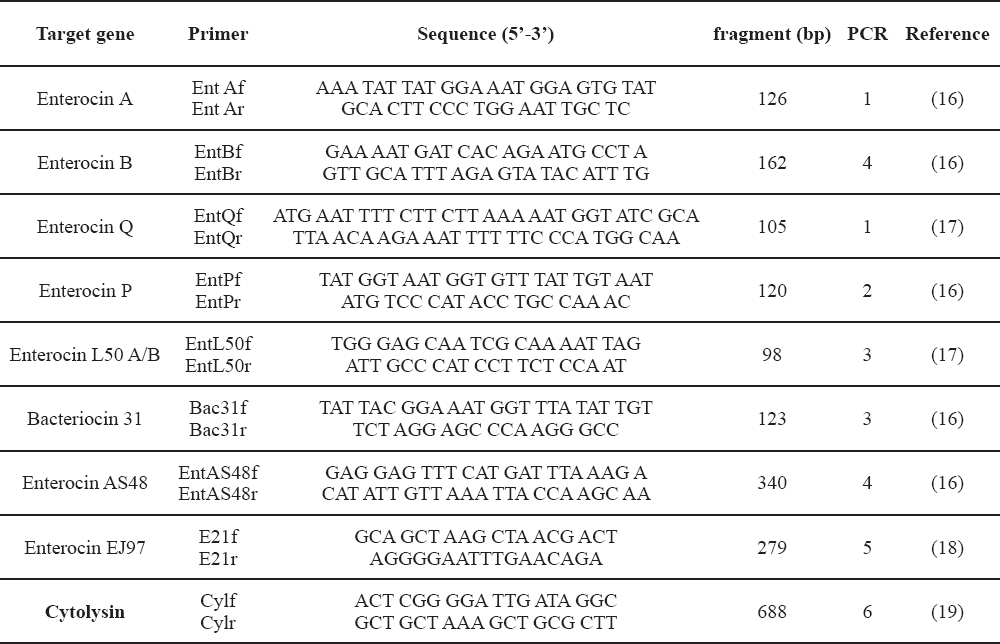
All isolates have been tested for the presence of the following bacteriocin structural genes: enterocin A (entA), enterocin B (entB), enterocin P (entP), enterocin L50A/B (entL50A-entL50B), bacteriocin 31 (bac31), enterocin AS48 (entAS48), enterocin Q (entQ), enterocin EJ97 (E21) and cytolysin (cyl) using specific enterocin PCR primers (Table 1).
Table 1. Primers used for PCR detection of bacteriocin genes
Primers have been used in combinations giving four different duplex and two single-plex PCR reactions, as described in Table 1 and Table 2. All PCR reactions were set in a final volume of 20 μl, consisting of 17 μl PCR mixture and 3 μl DNA template, using Taq PCR Mastermix Kit (Qiagen, USA). Positive controls were included in PCR assays for detection of EntA, EntB, EntP, E21, EntAS48 and cyl genes, whereas positive controls weren’t available for EntQ, Ent1071, Bac31, EntL50. Control DNA was obtained from isolates (FIFL-20, FIFL-36, FIFL-29, FIVRE-19) belonging to the culture collection of the Food Institute in which the presence of above mentioned structural genes has been confirmed by PCR.
Table 2. Enterocin genes and antimicrobial activity towards pathogens and their inhibition
The thermal profiles used for the amplification were the following: initial denaturation at 94° C for 3 minutes followed by 40 cycles of denaturation at 94 °C for 40 seconds, annealing (50° C for entA and ent Q and 56° C for entP, entB, ent50A/B, bac31, entAS-48, ent1071A/B, E21) for 40 seconds, and elongation at 72° C for 60 seconds. Final extension was at 72° C for 10 minutes. The thermal profile for cytolysin was the following: initial denaturation at 95° C for 2 minutes followed by 40 cycles of denaturation at 95 °C for 30 seconds, annealing at 56 °C for 90 seconds, elongation 72 °C for 90 seconds and final extension was at 72° C for 10 minutes. The amplification products were analysed by electrophoresis in 2.5% agarose gel at 7 V/cm for 1.5 h in Tris-acetate-EDTA buffer and revealed in ethidium bromide (20 µg/mL). The gel was photographed using UV light.
Bacteriocin-producer isolates were identified as E. faecalis (n=8), E. faecium (n=2) and E. hirae (n=2). Species determination for one of the isolates (E.spp.-90) was inconclusive: despite the positive identification using 16S rRNA PCR, BLAST analysis of the amplified part of 16s RNA sequence revealed 100% similarity with Lactococcus lactis subsp. lactis. This finding imposes the necessity for further and deeper analysis of this isolate.
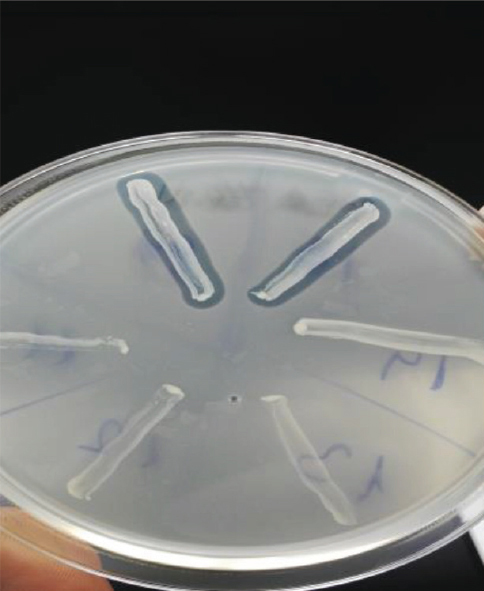
The 13 isolates were screened for antibacterial activity, since they may produce enterocins that can control the growth of the tested indicator bacteria. All of the isolates showed the activity predominantly against L. monocytogenes (Fig. 1), mostly showing larger inhibition zone for L. monocytogenes than L. innocua.
Figure 1 Inhibition areas against L. monocytogenes as a result of bacteriocin activity
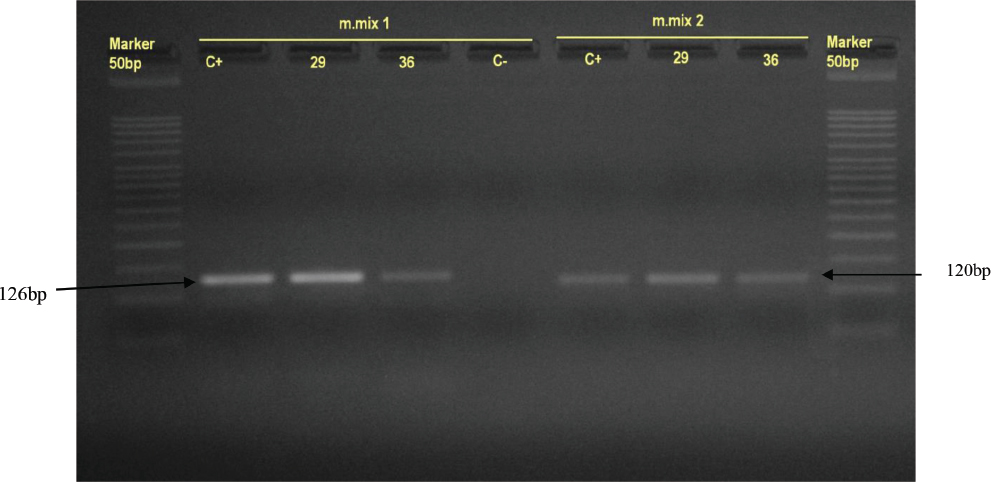
Figure 2 Agar gel visualisation of PCR products obtained with two duplex PCR reactions (PCR1 and PCR2). Lane Marker 50bp is 50 bp ladder, lanes C+ positive controls and lanes C- negative controls. Tested samples (29, 36) are in the lanes between positive and negative controls
Three of the isolates (E. faecalis-31, E.faecalis-32, E. hirae-36) were found to be inhibitory against S. aureus and P. aeruginosa, but only two of them were found to be inhibitory against B. cereus (E. faecalis-31 and E. hirae-36). None of the isolates were found to be inhibitory against S. enteritidis, Y. enterocolitica and E. coli (Table 2).
The most frequent bacteriocin structural genes in this study were enterocin P, enterocin A and cytolysin, followed by enterocin B. Less frequent genes were EJ97 and AS-48. Enterocin Q, bacteriocin 31 and enterocin L50 were not found in the tested enterococci (Table 2). E. faecalis-4, E.faecium-15, E. faecium-29 and E. hirae-36 had three different structural enterocin genes. E. faecalis-31, E. faecalis-32, E. faecalis-51, E. faecalis-77 and E. spp. had only one structural enterocin gene.
Two of the isolates (E. faecalis -25 and E. faecalis-86), did not harbour any of the tested bacteriocins.
A gene for cytolysin was detected in 5 isolates of E. faecalis (E. faecalis-4, E. faecalis-8, E. faecalis-31, E. faecalis-32, E. faecalis -51).
Results from previous studies report identical findings that most enterocin producing enterococci show antilisterial activity, while a small part of them also show inhibitory activity against Staphylococcus aureus and Bacillus spp. (7, 20, 21). Gram-negative bacteria are mostly resistant to many enterocins and the absence of inhibitory activity against them is in agreement with the data that most of the bacteriocins produced by Enterococcus strains inhibit the growth of closely related bacteria and activity against Gram-negative bacteria is very rare (22, 23). Though, we found inhibitory activity against Pseudomonas aeruginosa. There are reports that certain lactic acid bacteria especially the class 2 bacteriocins - pediocins can inhibit a limited number of Gram-negative bacteria including Pseudomonas, Shigella sp. Salmonella sp. (24, 25).
According to our PCR results, different bacteriocin structural genes are spread among enterococcal strains. Enterocin A, B and P were the most prevalent among enterococci which is in accordance with the results obtained for enterococci isolated from different sources of food and feed, animal isolates, clinical and nonclinical human isolates (26), enterococci isolated from artisan food of animal origin (27) and Spanish goats milk cheeses (28). Earlier study showed that entAS-48 was widely distributed throughout E. faecalis (29), but there are also authors who did not detect the presence of enterocin AS-48 (27, 28). Contrary to our findings, no equivalence in results was found with this authors. Only in two isolates (E. faecalis-25 and E. faecalis-86), we could not detect presence of any of the tested enterocin genes. Still, these isolates showed inhibitory activity against. L. monocytogenes and L. innocua. This could be explained with the possible presence of other already known bacteriocins (non-tested in this study) or possible existence of novel bacteriocins. As previously published by Ozdemir et al., (30), bacteriocin production is correlated to the species. However, contrary to their findings we found in our study E. hirae and E. faecalis as bacteriocin producers.
Enterocin A and P belong to the family of pediocins and they are grouped as Class II 1, which are effective mostly against listeria (31). Over the past ten years, class II bacteriocins produced by LAB have brought a significant attention to their potential application in food industry as natural preservatives and in the medical sector as antibiotic supplements or antiviral agents (7, 32). Enterocin B belongs to the subgroup II 3, which are part of the non-pediocin type enterocin. Enterocin AS-48 belongs to the class of cyclic antibacterial peptides (33). It should be underlined that only one of our isolates (E. faecalis-77) showed exclusively the entAS-48 gene, which is opposite to the previous findings in the literature that this enterocin is widely distributed among E. faecalis and E. faecium (29).
The combination of three different enterocin genes was observed in four strains (E. faecalis-4, E. faecium-15, E. faecium-29, E. hirae-36). Combination of two enterocin genes was observed in two strains (E. faecalis-8, E. hirae-52). According to the reviewed literature enterocin B is found predominantly with enterocin A, considering the fact that transport genes for enterocin B producers are not found (26, 34). In this study enterocin B was found with enterocin A, but it was also found in combination with enterocin P. It is considered by some authors that the reason for the high incidence of enterocin A, B and P genes in enterococcal bacteriocinogenic strains that are isolated from food is not only to antagonize bacteria but also to reinforce the competition of the selected strains against the competitors of the same species (35). Another combination of two genes is enterocin P and EJ97 and enterocin P, enterocin A and enterocin EJ97. These combinations, which were not found in the previous studies, were found in two of our isolates (E. hirae-36 and E. hirae-52).
Enterocin EJ97 belongs to enterocins synthesized without a leader peptide that is active against Gram-positive bacteria as well as enterococci, several species of Bacillus, Listeria and strains of S.aureus (36). Even though both strains (E. hirae -36 and E. hirae-52) posses the same structural gene EJ97, E. hirae-36 had a larger zone of inhibition. This could be explained by two possible hypotheses: the antimicrobial activity was reinforced by the presence of enterocin P and enterocin A or eventual presence of “silent” gene(s) in E. hirae-52. Various authors report the presence of bacteriocin silent genes in enterococci (26, 37, 38).
Cytolysin was found only in E. faecalis isolates, which is the species with the highest occurrence of virulence genes. According to our results, it can be concluded that there is no large zone of inhibition around the strain that harbour a gene for cytolysin. Cytolysin is the only two-peptide lantiobiotic isolated from genus Enterococcus with cytolitic activity. Cytolysin is not considered useful due to the fact that it’s a virulence factor (39), so these strains are not safe for application.
Results of this study showed that further analysis of certain strains as adjunct cultures for the process of fermentation of dairy products is needed. Also further studies should be conducted in order to identify new antimicrobial substances produced by the enterococcal strains.
Due to the fact that cytolysin’s virulence factor is not considered useful, so these strains are not safe for application. The usefulness of the strains that are free of cytolysin genes should be investigated, because these strains could be candidates for safe and practical use. Having in mind the potential of the strains, the future challenge will be optimizing the conditions during the fermentation process, as well as the composition of the medium in order to achieve better production of bacteriocin substances.
As a conclusion, our investigation gives a clear evidence of bacteriocin producing enterococci in traditional artisanal-produced cheese. The main bacteriocinogenic species was E. faecalis. Enterocin structural genes can be found in many different combinations, some of them reported for the first time in this study.
Copyright
© 2015 Mojsova S. This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial License (http://creativecommons.org), which permits unrestricted non-commercial use, distribution, and reproduction in any medium provided the original work is properly cited.
Conflict of Interest Statement
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Citation Information
Macedonian Veterinary Review. Volume 38, Issue 2, Pages 209-216, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.14432/j.macvetrev.2015.08.052, 2015