Mac Vet Rev 2016; 39 (2): 175 - 183
 10.1515/macvetrev-2016-0083
10.1515/macvetrev-2016-0083
Received: 22 January 2016
Received in revised form: 25 April 2016
Accepted: 30 April 2016
Available Online First: 05 May 2016
Published on: 15 October 2016
Keywords: polyunsaturated fatty acids, ultrasound guided transvaginal follicular aspiration, cows, follicular growth
In the last decades, an intensive genetic selection for higher milk production has inevitably contributed to declining fertility in dairy cows (1). Several studies have shown a strong negative relationship between high milk production and fertility (2, 3) particularly during the period of negative energy balance. The latter occurs in the early postpartum period due to the incapability of the cows to meet their energy needs (4). Implementation of dietary fats in the daily ration in an effort to ameliorate negative energy balance could increase (at least part of) cow’s energy demands, leading to enhanced reproductive performance (5, 6). However, it was suggested that specific fatty acids (FA), independent of energy status, might be involved and could affect follicle diameter (7) corpus luteum (CL) function (8), oocyte and embryo quality (9), steroidogenesis (10) and prostaglandin synthesis (11). In this respect, the two main families of polyunsaturated fatty acids (PUFA), omega-3 (n-3, linolenic acid, C18:3n3) and omega-6 (n-6, linoleic acid, C18:2n6) were reported to affect fertility (12), although there is some inconsistency in the published literature on this topic (13).
The composition of the follicular fluid (FF) might be of great importance in ovarian physiology, including steroidogenesis, growth of the follicle and maturation of the oocyte and ovulation (14). Follicular fluid is composed partly of secretions from granulosa and thecal cells, and partly of constituents from plasma that cross the blood follicular barrier (14, 15). Therefore, any alterations in the components of the plasma and granulosa and theca layers will be reflected in the follicular fluid (16). In this respect, it has been shown that alterations in the diet that increase the serum FA, especially PUFA levels, cause considerably greater infiltration of these FA into the follicular fluid (17). Increased follicular fluid PUFA concentrations might increase the pre-ovulatory follicular diameter (11, 16) which could result in the subsequent formation of a larger CL capable to produce greater amounts of progesterone (18). Furthermore, increased PUFA concentrations (C18:3n3) have been shown to decrease PGF2α synthesis (19), which can prolong the lifespan of the CL; both effects being reported to be beneficial for embryo survival (20). However, some authors reported that the FA composition in the FF is different from that in the plasma and that the former depends mostly on the estrogenic activity of the follicles (21). Indeed, it has been demonstrated that follicles have their own unique lipid metabolism and FA composition, the latter not directly reflecting the FA profile of the serum (22). Therefore, changes in the FA composition either in the blood or in the follicular fluid may have a pivotal role in many reproductive responses and could affect subsequent fertility.The precise mechanism how they mediate the positive effects on the reproductive performance in the dairy cows, however, warrants further investigation.
The use of PGF2α to synchronize estrus is the most common protocol implemented in dairy farms. An intramuscular injection of PGF2α or its analogues to cows with a functionally mature corpus luteum (CL), in 85% to 95% of the cases leads to luteolysis followed by the initiation of a new follicular phase (23). However, the interval from treatment to ovulation differs between treated animals due to variation in the size of the follicle present at the moment of PGF2α application (24). Recently, we have shown that cows treated with PGF2α when no dominant follicle is present have a numerically faster daily growth rate, and need a significantly longer interval to attain ovulatory follicles, in comparison to cows that already have follicles larger than 8.5 mm at the time of treatment (25).
We hypothesized that the FA content in the blood and/or in the FF might be associated with the follicular dynamics after PGF2α injection. Therefore, the objectives of the present study were to examine 1) the FA profiles in the serum and in the FF and 2) relations between PUFA and follicular growth dynamics after induced luteolysis using a single PGF2α application in cows fed a standard lactation ration without any fat supplementation.
A total of 29 Holstein-Friesian dairy cows from two dairy farms in the Republic of Macedonia were included in the study. In Dairy 1 (n=15), cows were housed in a free stall with cubicles, whereas in Dairy 2 (n=14) cows were housed in a tie-stall on deep straw bedding. On both farms, cows were fed a standard TMR based on corn silage, chopped alfalfa, straw and a concentrate-mineral mix offered twice daily. On Dairy 1 and 2, cows were milked twice or thrice daily with an average 305 d-milk production level of 6400 and 6500 kg, respectively. Cows fulfilling the following criteria were included in the study: displaying a regular ovarian cyclicity (regular cycles of approximately 21 days), and at d0 (start of the experiment), presence of a mature CL (> 25 mm) and a follicle ≈15 mm. At d0, all cows were submitted to an ultrasound-guided trans-vaginal follicle aspiration (UTFA) for collection of FF from the largest follicle (defined as the follicle exceeding the size of other follicles on both ovaries). The same day, all animals received 500 μg of cloprostenol i.m to induce luteolysis. According to the follicular growth patterns following aspiration, the cows were subdivided into three groups: Group A1 (n=11) representing the cows which resumed follicular growth from follicles less than 8.5 mm, Group A2 (n=8) cows resuming follicular growth from the second largest follicle present at the moment of aspiration (follicle larger than 8.5 mm) and Group A0 (n=10) cows that did not resume follicular growth after aspiration. Ovarian follicular development was monitored daily by transrectal ultrasonography starting at d0 until the subsequent dominant follicle reached the size of ≈ 15 mm. When, in the cows that resumed regular follicular growth the subsequent dominant follicle reached the size of ≈ 15 mm (referred hereafter as d1), another UTFA was performed for FF collection. Follicular fluid samples were immediately refrigerated at +4° and transported within 3 hours to the laboratory for further analysis. In total, each group represented a specific developmental stage of the dominant follicle at the moment of the injection of a luteolytic dose of prostaglandins: no follicular growth (Group A0), no dominant follicle <8.5mm (Group A1) and presence of a dominant follicle >8.5mm (Group A2). Blood samples for serum FA determination were taken on d0 in all cows and simultaneously with FF collection in Group A1 and A2 on d1 from the coccygeal vein into glass tubes and immediately refrigerated at +4°. Serum was obtained at 4000 RPM, 10 min at +4° within 3 hours and frozen at -18° until analysis.
Ultrasonographic examinations were performed using a B-mode scanner Aloka SSD 500 (Tokyo, Japan), equipped with a 7.5 MHz linear-array transducer for intra-rectal use (ultrasonographic examination of the ovaries) and convex-array transducer (5 MHz) for intra-vaginal use (UTFA). Before insertion of the lubricated transducer, the rectum was emptied, and the ovaries were first manually located before introducing the transducer. Diameters of the follicles were measured by means of electronic calipers located on the ultrasound device after freezing the pictures on the screen. The daily follicular growth rates and luteolysis were estimated in all groups using the definitions as described in the concomitant study (25). Briefly, daily follicular growth rate (cm/day) - the difference between the first visualization of the follicle and 15 mm divided by the number of days necessary to attain 15 mm (modified by Kojima et al. (26)). Luteolysis - (structural and functional regression of the CL), decreasing of the CL diameter concomitantly with a decline in P4 concentration below 0.5 ng/mL (modified from Martins et al., (27)).
Collection of the FF was done using UTFA as described by Bergfelt et al. (28). Briefly, caudal epidural anesthesia was performed using 3 to 6 ml of a 2% lidocaine solution (Rotexmedicca, Trittau – Germany). Next, the perineal area was thoroughly cleansed and disinfected using 10% povidone-iodine solution (Betadine, Alkaloid, R. Macedonia). A convex-array 5 MHZ transducer was placed in the anterior vagina; it was equipped with a needle guide for guiding the aspiration needle through the vaginal wall into the ovaries. While the transducer was positioned in the vaginal fornix, the free hand was used transrectally to position the ovary, against the vaginal wall. The follicle was stabilized over the transducer face. A 17-gauge, single lumen needle (55 cm long) was placed in the needle guide and advanced through the vaginal wall and into the follicular antrum. Ultrasonic hyper-echogenicity at the needle tip, as well as a guide-line on the screen of the ultrasound device allowed determination of its location and direction during follicle puncture. Follicular fluid was manually aspirated by applying a vacuum using a 10 ml plastic syringe settled upon the opposite side of the needle.
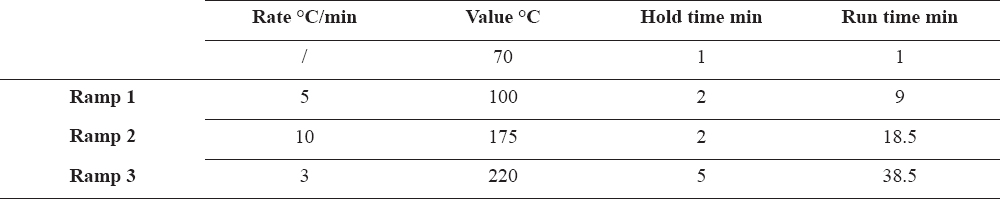
The lipids from the serum and follicular fluid were extracted according to the modified AOAC Official Method 996.06 (29) and methylation of the samples was performed with the same method. Determinations of the FA-methyl esters were carried out at the Laboratory for residues and contaminants at the Faculty of Veterinary Medicine – Skopje, on a gas chromatography with flame ionization detector (GC-FID, GC Agilent Technologies 7890 GC System, USA). The column HP88 from Agilent (J&W HP88 capillary column 112 -8867, 60m x 250mm x 0.2 mm) was used. The operation parameters of the column are given in Table 1.
Table 1. Operation parameters of the capillary column
The injector was kept at 250 ºC and the detector at 300 ºC. The gas flows on the GC-FID system were: 1.4 mL/min carrier gas (He), 23 mL/min make up gas (N2), 30 mL/min H2 and 400 mL/min flame synthetic air. The ratio used was 200:1 and the injection volume was 1 μL. Fatty acid methyl esters (FAMEs) were identified by comparing the sample FAME peak retention times and peak area with those obtained for a standard FAME mix (Supelco 37 component FAME mix, Sigma-Aldrich, Germany). The method was verified according to the guidelines for validation of chromatographic methods (30).
All statistical analyses were performed using the SAS software package (Statistical Analyses System Institute, Inc., Cary, NC, USA, 2010). Descriptive statistics were done using PROC MEANS and PROC FREQ. Normality of the variables was checked using the Kolmogorov-Smirnov method. Correlations between variables were checked using PROC CORR. The MIXED procedure for repeated measures was used to analyze the effect of groups (A0, A1 and A2), day of puncture (d0 and d1), and luteolysis (yes and no) on the FA composition, except for C18:3n3 for which the GLIMMIX procedure with lognormal distribution was used. Data are reported as model least square means with standard errors (LS means ± SEM) unless indicated otherwise. Significance and tendency were declared at p< 0.05 and 0.05 <p< 0.1 respectively.
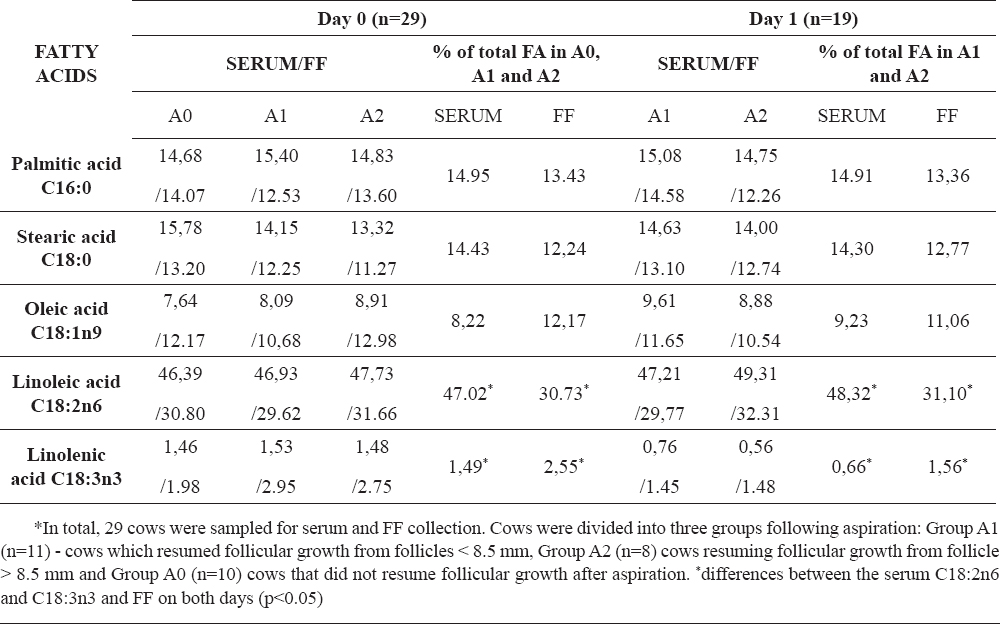
The serum and FF FA are shown in Table 2. In all cows, the FA with the highest abundance (percentage) found in the serum was linoleic acid, followed by respectively palmitic, stearic and oleic acid. Similarly, the most abundant FA in the FF was linoleic acid, followed by respectively palmitic, stearic and oleic acid. No differences were observed in the FA profiles in the serum and in the FF between both days (p>0.05). Additionally, the FA profile in the serum was not significantly different from the FA profile in the FF between the cows in all groups on d0 and between the cows on d1 (A1 and A2).
Table 2. Serum and follicular fluid (FF) fatty acids in all cows (% of total fatty acids)
In respect of C18:2n6, the serum percentages of C18:2n6 at d0 (47.02 ± 3.56) and d1 (48.32 ± 3.01) were significantly higher than the percentage in the FF on the same days (d0, 30.73 ± 6.91 and d1, 31.10 ± 7.55, p<0.05, respectively) in the cows from all groups. However, on both days no differences were observed between C18:2n6 neither in the serum nor in the FF between the groups (p>0.05). Additionally, the serum C18:2n6 percentages were negatively correlated with the remaining serum FAs: C18:0 (r= 0.42, p<0.05), C16:0 (r= 0.30, p<0.05), C18:1 (r= 0.27, p=0.05), and C18:3 (r= 0.34, p<0.01), respectively, whereas FF C18:2n6 percentages were negatively correlated with the C18:1 (r= 0.39, p<0.05) on both days.
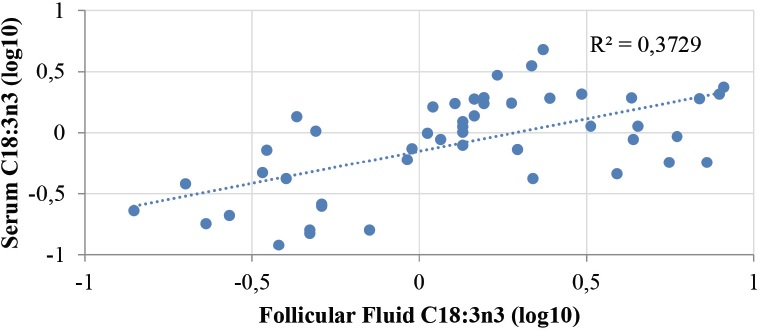
Regarding the C18:3n3, there was a tendency in cows of all groups to have a lower percentage of C18:3n3 in the serum than in the FF on both d0 and d1 (1.49 ± 1.00 vs.2.55 ± 2.15 p=0.08, and 0.66 ± 0.60 vs. 1.56 ± 2.11, p=0.05), respectively. However, no significant differences were observed in the serum C18:3n3 levels between the groups on both days (p>0.05). In addition, the percentage of C18:3n3 in the serum on d0 (1.49 ± 1.00) was significantly higher (p<0.05) than the percentage on d1 (0.66 ± 0.60) in all cows. Similarly, the percentage of C18:3n3 in the FF was significantly higher on d0 than d1 (2.55 ± 2.15 vs. 1.56 ± 2.11, p<0.05), respectively. However, no differences were observed between the groups on both days (p>0.05). Furthermore, there was a strong positive correlation between the serum and FF C18:3n3, r=0.61, p<0.0001 on both days (Fig. 1). Additionally, despite the previously observed negative correlation with C18:2n6 (see above) in the serum, the serum and FF C18:3n3 levels were not significantly correlated with the remaining serum and FF FA concentrations.
Figure 1. Correlation between C18:3n3 in the serum and in the follicular fluid (r=0.61, p<0.0001)
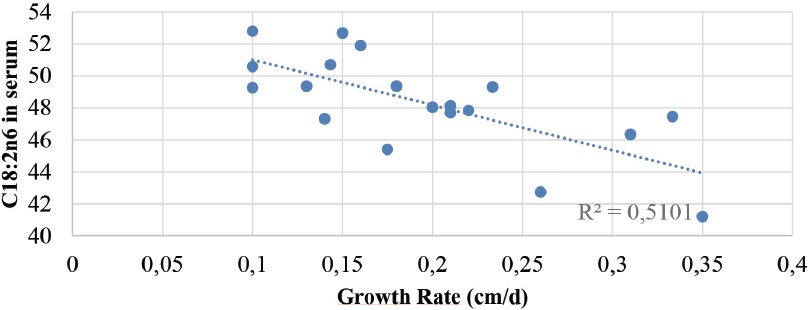
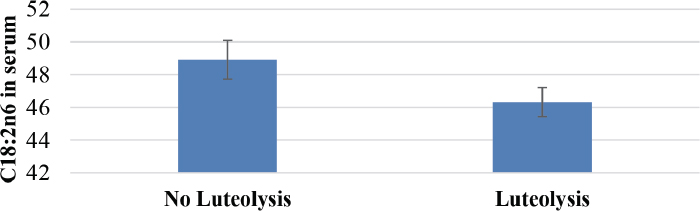
On d0, there was a tendency of both the serum and the FF C18:2n6 to be negatively correlated with the daily follicular growth rate (r= 0.44, p=0.05 and, r= 0.39, p=0.09, respectively). On d1, a strong negative correlation (Fig. 2) between the serum C18:2n6 and the daily growth rate was observed (r= 0.71, p=0.0006). On the contrary, the serum and FF C18:3n3 levels were not correlated with the daily follicular growth rate. Additionally, the serum C18:2n6 tended to have an effect on luteolysis (p=0.06, Fig. 3).
Figure 2. Negative correlation between the serums C18:2n6 concentration and the daily follicular growth rate (r=-0.71, p=0.0006)
Figure 3. Effect of the serum C18:2n-6 concentration on the occurrence of luteolysis
The current study aimed to examine the correlation between FA profiles in the serum and the follicular fluid in cows fed a lactation diet without any fat supplementation and to investigate if the PUFA fatty acid concentrations (C18:2n6 and C18:3n3) are associated with the follicular growth dynamics in dairy cattle after a single PGF2α injection.
The results of the present study show that linoleic acid (C18:2n6) was the principal fatty acid constituting about half of the total amount of fatty acids in the serum and about a third of the total fatty acid concentration in the follicular fluid followed by palmitic acid, stearic acid and oleic acid, respectively. Similarly, one study (22) reported that the same fatty acids were present in the serum as well as in the follicular fluid with the highest concentrations of linoleic acid, followed by oleic acid, stearic acid and palmitic acid. Moreover, our results confirm the results in the literature (31, 32) in which C18:2n6 was the predominant FA in the serum as well as the most abundant FA in the follicular fluid constituting about either half or a third of the total fatty acids, respectively. As a contrast to our results were the results reported by Khandoker et al. (33) showing that the predominant fatty acid in the follicular fluid was oleic acid, followed by palmitic, stearic and linoleic acid. Nevertheless, when comparison in the present study was made between the serum and the follicular fluid FA profiles, no significant differences were revealed. The latter is in agreement with the literature (34) indicating that FAs profiles of the FF are very similar to those in the serum. Since follicular fluid obtains metabolites partly from the serum (14), any dietary alterations that are effectively transferred in the serum may modify the fatty-acid composition of the follicular fluid, which, in its turn can be crucial for altering ovarian activity and oocyte environment (17, 35). In this respect, incorporations of PUFA into the rations might improve fertility, since, it has been reported that in general, unsaturated fatty acids tend to have beneficial effects when present in elevated concentrations on early embryo development (36). However, in contrast to our findings, other studies have reported (21, 22) that the follicular fluid has a unique lipid composition which does not represent a simple reflection of the serum but significantly depends on the estrogen activity of the follicles. Since the UTFA of the follicles, in the present study, was done simultaneously with the collection of the blood samples, observations of similar FA profiles between serum and FF, additionally, do not support the findings observed in those studies (21, 22).
Furthermore, our results show that linoleic acid either in the serum or in the follicular fluid was not different between the groups. However, the serum C18:2n6 was significantly higher than in the follicular fluid. The latter is in agreement with the results observed by Aardema et al. (37) showing that a short elevation of C18:2n6 in the serum did not result in a similar shift in the FF. These findings indicate the existence of a selective mechanism in the follicle that regulates the content of the C18:2n6 concentration in the follicular fluid being particularly important for the developmental competence of the oocyte, since the FF C18:2n6 concentration has been shown to significantly inhibit the resumption of meiosis (32). Additionally, in the present study, a negative correlation of both serum and FF C18:2n6 with the daily follicular growth rate was observed. Currently, there is a lack of data in the literature to clarify all ambivalent results reported in the different studies, which necessitates further investigations. Furthermore, the serum C18:2n6 concentration showed a tendency to have an effect on the occurrence of luteolysis, which is in agreement with the literature (31), since linoleic acid (by its desaturation and elongation) can be converted to arachidonic acid (C20:4n6) a precursor of PGF2α (19, 38), a natural luteolysin. This suggests that feeding the cows diets enriched with linoleic acid, allows a greater pool of linoleic acid and increased substrate concentrations of arachidonic acid which might subsequently increase PGF2α synthesis and cause premature luteal regression leading to decreased pregnancy results. We speculate that the latter might be a reason of the lower percentage of C18:2n6 in the FF observed in the current study.
Our results additionally demonstrate that cows in all groups tend to have a lower serum linolenic acid (C18:3n3) concentration than in the follicular fluid, indicating that as the follicle starts or continues to grow, the serum C18:3n3 fatty acid is effectively transferred into the follicular fluid which might subsequently influence the follicular status. Similarly, Bender et al. (22) observed significant differences between dominant versus subordinate follicles, in the content of C18:3n3 in the follicular fluid, signifying that its concentration is increasing in the dominant follicle after reaching dominancy. Since an increased C18:3n3 concentration might influence the dominant follicle diameter (7), which acts as the primary determinant of the treatment-estrus interval in a single PGF2α synchronization regime (24), it can be assumed that elevated C18:3n3 in the follicular fluid might affect the follicular dynamics (7). Indeed, some authors (17) have shown that the effects on ovarian function are related to the modification in fatty acid composition, however, we were not able to observe that the C18:3n3 in the FF is related with the daily follicular growth rate. Although, in the present study, the actual concentrations of the C18:3n3 in the FF was not estimated, we speculate that the lower observed percentage of the C18:3n3 in FF might be insufficient to affect the follicular growth. Additionally, in the current study, there was a very strong positive correlation between the serum and the follicular fluid C18:3n3 fatty acid concentration. Surprisingly, in our previous findings (39), we observed a negative correlation for the percentage of C18:3n3 (r= 0.47) between the follicular fluid and the serum. The reason for the inconsistent results still remains unclear, although in general, the latter might lead to the assumption that follicles indeed posses a selective up-take mechanism against major fluctuations of plasma PUFA which was also reported to exist (40), providing optimal environment for the oocyte development.
Feeding dairy cows a daily ration without any fat supplementation, results in similar fatty acid profiles in the serum and in the follicular fluid. The major constituent in the serum and in the follicular fluid is linoleic acid (C18:2n6) which has been shown to be negatively correlated with the daily follicular growth and might affect the occurrence of luteolysis after a single PGF2α administration. On the contrary, linolenic acid (C18:3n3) was not associated with follicular growth despite its positive correlation between the serum and the follicular fluid. Therefore, it can be assumed that alteration of the serum PUFA profile may cause a similar modification to the PUFA content in the follicular fluid, which might influence the ovarian dynamics and subsequently the fertility. However, the exact mechanisms by which PUFA have an impact on the follicular growth, warrant further investigation.
1. Lucy M.C, Reproductive loss in high-producing dairy cattle: where will it end?J. Dairy Sci 2001; 84: 1277-1293. http://dx.doi.org/10.3168/jds.S0022-0302(01)70158-0
2. Royal M.D, Darwash A.O, Flint A.P.F, Webb R, Woolliams J.A, Lamming G.E, Declining fertility in dairy cattle: changes in traditional and endocrine parameters of fertilityAnim. Sci 2000; 70: 487-501.
3. Washburn S.P, Silvia W.J, Brown C.H, McDaniel B.T, McAllister A.J, Trends in reproductive performance in southeastern Holstein and Jersey DHI herdsJ. Dairy Sci 2002; 85: 244-251. http://dx.doi.org/10.3168/jds.S0022-0302(02)74073-3
4. Santos J.E.P, Reproductive management of lactating dairy cows for first postpartum inseminationDairy production medicine 2011; John Wiley & Sons Inc; 81-98.
http://dx.doi.org/10.1002/9780470960554.ch7
5. Lucy M.C, De La Sota R.L, Staples C.R, Thatcher W.W, Effect of dietary calcium salts of long chain fatty acids (CaLCFA), energy intake, and lactation on ovarian follicular dynamics in Holstein dairy cowsJ. Anim. Sci 199la; 69: 1451-Abstr
6. Ambrose D.J, Kastelic J P, Dietary fatty acids and dairy cow fertilityAdvances in Dairy Technology 2003; 15: 35-47.
7. Ambrose D.J, Kastelic J.P, Corbett R, Pitney P.A, Petit H.V, Small J.A, Zalkovic P, Lower pregnancy losses in lactating dairy cows fed a diet enriched in α-linolenic acidJ. Dairy Sci 2006; 89: 3066-3074. http://dx.doi.org/10.3168/jds.S0022-0302(06)72581-4
8. Petit H.V, Dewhurst R.J, Scollan N.D, Proulx J.G, Khalid M, Haresign W, Twagiramungu H, Mann G.E, Milk production and composition, ovarian function, and prostaglandin secretion of dairy cows fed Omega-3 fatsJ. Dairy Sci 2002; 85: 889-899. http://dx.doi.org/10.3168/jds.S0022-0302(02)74147-7
9. Fouladi-Nashta A.A, Gutierrez C.G, Gong J.G, Garnsworthy P.C, Webb R, Impact of dietary fatty acids on oocyte quality and development in lactating dairy cowsBio of Repro 2007; 77: 9-17. http://dx.doi.org/10.1095/biolreprod.106.058578 PMid: 17344470
10. Robinson R.S, Pushpakumara P.G, Cheng Z, Peters A.R, Abayasekara D.R, Wathes D.C, Effects of dietary polyunsaturated fatty acids on ovarian and uterine function in lactating dairy cowsReproduction 2002; 124: 119-131. http://dx.doi.org/10.1530/rep.0.1240119
11. Abayasekara D.R, Wathes D.C, Effects of altering dietary fatty acid composition on prostaglandin synthesis and fertilityProstaglandins, Leukotrienes and Essential Fatty Acids 1999; 61: 275-287. http://dx.doi.org/10.1054/plef.1999.0101 PMid: 10670689
12. Staples C.R, Thatcher W.W, Nutrient influences on reproduction of dairy cowsMid-South Ruminant Nutrition Conference 2001; 21-36.
13. Gulliver C.E, Frienda M.A, Kinga B.J, Claytonc E.H, The role of omega-3 polyunsaturated fatty acids in reproduction of sheep and cattleAnim. Repro Sci 2012; 131: 9-22. http://dx.doi.org/10.1016/j.anireprosci.2012.02.002 PMid: 22386690
14. Edwards R.G, Follicular fluidJ Reprod Fertil 1974; 37: 189-219. http://dx.doi.org/10.1530/jrf.0.0370189
15. Gosden R.G, Hunter R.H, Telfer E, Torrance C, Brown N, Physiological factors underlying the formation of ovarian follicular fluidJ Reprod Fertil 1988; 82: 813-825.
http://dx.doi.org/10.1530/jrf.0.0820813
16. Zachut M, Arieli A, Lehrer H, Argov N, Moallem U, Dietary unsaturated fatty acids influence preovulatory follicle characteristics in dairy cowsReproduction 2008; 135: 683-692. http://dx.doi.org/10.1530/REP-07-0556 PMid: 18296508
17. Zachut M, Dekel I, Lehrer H, Arieli A, Arav A, Livshitz L, Yakoby S, Moallem U, Effects of dietary fats differing in n-6: n-3 ratio fed to high-yielding dairy cows on fatty acid composition of ovarian compartments, follicular status, and oocyte qualityJ. Dairy Sci 2010; 93: 529-545. http://dx.doi.org/10.3168/jds.2009-2167 PMid: 20105525
18. Pfeifer L.F.M, Mapletoft R.J, Kastelic J.P, Effects of low versus physiologic plasma progesterone concentrations on ovarian follicular development and fertility in beef cattleTheriogenology 2009; 72: 1237-1250. http://dx.doi.org/10.1016/j.theriogenology.2009.07.019 PMid: 19781753
19. Ricardo M, Staples C.R, Thatcher W.W, Effects of dietary fatty acids on reproduction in ruminantsReviews of Reproduction 2000; 5: 38-45. http://dx.doi.org/10.1530/ror.0.0050038
20. Wiltbank M.C, Souza A.H, Carvalho P.D, Cunha A.P, Giordano J.O, Fricke P.M, Baez G.M, Diskin M.G, Physiological and practical effects of progesterone on reproduction in dairy cattleThe Animal Consortium 2014; 1-12. http://dx.doi.org/10.1017/s1751731114000585
21. Renaville B, Bacciu N, Comin A, Motta M, Poli I, Vanini G, Prandi A, Plasma and follicular fluid fatty acid profiles in dairy cowsRepro. Dom. Anim 2010; 45: 118-121.
http://dx.doi.org/10.1111/j.1439-0531.2008.01264.x PMid: 19055554
22. Bender K, Walsh S, Evans A.C.O, Fairm T, Brennan L, Metabolite concentrations in follicular fluid may explain differences in fertility between heifers and lactating cowsReproduction 2010; 139: 1047-1055. http://dx.doi.org/10.1530/REP-10-0068 PMid: 20385782
23. MacMillan K.L, Prostaglandin responses in dairy herd breeding programN Z Vet. J 1983; 31: 110-114. http://dx.doi.org/10.1080/00480169.1983.34987 PMid: 16030977
24. Kastelic J.P, Knopf L, Ginter O.J, Effect of day of prostaglandin F2αtreatment on selection and development of the follicle in heifersAnim Repro Sci 1990; 23: 169-180.
http://dx.doi.org/10.1016/0378-4320(90)90001-V
25. Atanasov B, Hostens M, Celeska I, Ilieska K, Opsomer G, Dovenski T, Follicular dynamics following induced luteolysis and transvaginal ultrasound-guided aspiration of the largest follicle in dairy cowsVet arhiv 2015; 85: 3247-260.
26. Kojima F.N, Bergelt E.G, Wehrman M.E, Cupp A.S, Fike K.E, Frequency of luteinizing hormone pulses in cattle influences duration of persistence of dominant ovarian follicles, follicular fluid concentration of steroids and activity of insulin-like growth factor binding proteinsAni. Repro. Sci 2003; 77: 187-211. http://dx.doi.org/10.1016/S0378-4320(03)00038-1
27. Martins J. P. N, Policelli R.K, Neuder L.M, Raphael W, Pursley J. R, Effects of cloprostenol sodium at final prostaglandin F2αof Ovsynch on complete luteolysis and pregnancy per artificial insemination in lactating dairy cows JDairy Sci 2011; 94: 2815-2824. http://dx.doi.org/10.3168/jds.2010-3652 PMid: 21605751
28. Bergfelt D.R, Lightfoot K.C, Adams G.P, Ovarian synchronization following ultrasound-guided transvaginal follicle ablation in heifersTheriogenology 1994; 42: 895-907. http://dx.doi.org/10.1016/0093-691X(94)90113-W
29. William H, Latimer G.W, AOAC Official Method 996.06, Fat (total, saturated and unsaturated) in Foods 2005;
30. Taverniers I, De Loose M, Van Bockstaele E, Trends in quality in the analytical laboratory: Analytical method validation and quality assuranceTrends in Analytical Chemistry 2004; 23: 535-552. http://dx.doi.org/10.1016/j.trac.2004.04.001
31. Petit H.V, Germiquet C, Lebel D, Effect of feeding whole, unprocessed sunflower seeds and flaxseed on milk production, milk composition, and prostaglandin secretion in dairy cowsJ. Dairy Sci 2004; 87: 3889-3898. http://dx.doi.org/10.3168/jds.S0022-0302(04)73528-6
32. Homa S.T, Brown C.A, Changes in linoleic acid during follicular development and inhibition of spontaneous breakdown of germinal vesicles in cumulus-free bovine oocytesJ. Repro. Fert 1992; 94: 153-160. http://dx.doi.org/10.1530/jrf.0.0940153
33. Khandoker M.A.M, Tsujii H, Karasawa D, Fatty acid composition of oocytes, follicular, oviductal and uterine fluids of pig and cowAsian-Aust. J. Anim. Sci 1997; 10: 523-527. http://dx.doi.org/10.5713/ajas.1997.523
34. Zachut M, Arieli A, Moallem U, Incorporation of dietary n-3 fatty acids into ovarian compartments in dairy cows and the effects on hormonal and behavioral patterns around estrusReproduction 2011; 141: 833-840. http://dx.doi.org/10.1530/REP-10-0518 PMid: 21389076
35. Moallem U, Shafran A, Zachut M, Dekel I, Portnick Y, Arieli A, Dietary α-linolenic acid from flaxseed oil improved folliculogenesis and IVF performance in dairy cows, similar to eicosapentaenoic and docosahexaenoic acids from fish oilReproduction 2013; 146: 603-614. http://dx.doi.org/10.1530/REP-13-0244 PMid: 24062566
36. Leroy JLMR, Sturmey R.G, Van Hoeck V, De Bie J, McKeegan P.J, Bols PE.J, Dietary fat supplementation and the consequences for oocyte and embryo quality: hype or significant benefit for dairy cow reproduction? ReproDom. Anim 2014; 49: 353-361. http://dx.doi.org/10.1111/rda.12308 PMid: 24697981
37. Aardema H. F, Lolicato C.H.A, van de Lest J.F, Brouwers A.B, Vaandrager H.T.A, van Tol B.A.J, Roelen PLAM, Vos J.B, Helms Gadella B.M, Bovine cumulus cells protect maturing oocytes from increased fatty acid levels by massive intracellular lipid storageBiol of Repro 2013; 88: 1641-15. http://dx.doi.org/10.1095/biolreprod.112.106062
38. Thatcher W, Santos J.E.P, Staples C.R, Dietary manipulations to improve embryonic survival in cattleTheriogenology 2011; 76: 1619-1631.
http://dx.doi.org/10.1016/j.theriogenology.2011.06.005 PMid: 21924473
39. Dovenski T, Trojacanec P, Hajrulai Musliu Z, Atanasov B, Uzunov R, Jasari B, Dovenska M, Grizelj J, Comparison between fatty acid composition of follicular fluid and blood serum in dairy cows with ovarian disordersProceeding of 4nd General Meeting of GEMINI Maternal communication with Gametes and Embryo 2011;
40. Marei W.F, Wathes D.C, Fouladi-Nashta A.A, The effect of linolenic acid on bovine oocyte maturation and developmentBiol of Repro 2009; 81: 1064-1072.
http://dx.doi.org/10.1095/biolreprod.109.076851 PMid: 19587335
© 2016 Atanasov B. This is an open-access article publishedunder the terms of the Creative Commons Attribution License whichpermits unrestricted use, distribution, and reproduction in any medium,provided the original author and source are credited.
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 39, Issue 2, Pages 175-183, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.1515/macvetrev-2016-0083, 2016