Mac Vet Rev 2017; 40 (1): 5 - 11
 10.1515/macvetrev-2016-0091
10.1515/macvetrev-2016-0091
Received: 02 March 2016
Received in revised form: 13 June 2016
Accepted: 20 September 2016
Available Online First: 24 October 2016
Published on: 15 March 2017
Keywords: Cyprinus, pseudoeosinophilic granulocytes, acclimation, stress
Different environmental stress factors affect the fish organism in intensive fish culture. Deleterious stimuli such as temperature changes, variation of pH, decrease in oxygen concentration, increase in ammonia concentration, handling, transport and osmotic changes in water induce stress reaction in fish (1, 2). A stress reaction, which is an integrated response with behavioral, neural, hormonal and physiological elements by the fish organism can cause changes in the fish health status and reduce the resistance to diseases (3, 4). Changes in water quality can lead to significant disturbances in fish, such as various diseases and very significant hematological changes in the blood of fish (5). Cellular stress response is a protective response of individual cells to potentially harmful stimuli from the environment (6). Cellular stress response was first described as a reaction to increased temperature (7). To counter stress, the cells increase the expression of chaperones that help in protein refolding and aggregation alleviating (8). The cell thus provides a short-term protection, leading to a condition called thermotolerance and cells become resistant to a variety of negative effects (9, 10). One of the important responses in fish adaptation is the hematological response, especially the number of leukocytes, and it is the first defense line against stress agents (9). The differentiation of the blood cells of fish is much debatable, but there is almost no uniform classification or reference interval, neither for fish families neither for the level of the genus Cyprinus (11). Fish blood is very rich in leukocytes, which are divided into granulocytes (neutrophils, eosinophils, basophils) and agranulocytes (lymphocytes and monocytes).
Pseudoeosinophilic granulocytes are known as heterophils (12). They are large round cells, with eccentric round to oval nucleus. Eosinophilic, basophilic and neutrophilic granulation are present in cytoplasm whose number depends on the degree of maturation of the cells. It is important to emphasize the simultaneous presence of granules that have different colors (13, 14). A significant increase of pseudoeosinophils marked as pseudoeosinophilia was recorded during stress (15). Most researches compare the functional significance of pseudoeosinophils with neutrophils despite their different morphological characteristics (16). It has been reported that pseudoeosinophils are functionally heterogeneous cells with combined morphological elements and the roles of other granulocytes. Recent researches show that pseudoeosinophilic granulocytes are present in wild fish, especially in tench (Tinca tinca Linnaeus, 1758) and sporadically in European chub (Squalius cephalus Linnaeus, 1758) (17). It is interesting that crucian carp (Carassius carassius Linnaeus, 1758) does not have pseudoeosinophilic granulocytes in their blood, nor does the rainbow trout (Oncorhynchus mykiss Walbaum, 1792) in the wild (18). Artificially bred trout does not have the ability for a longer survival rate in warmer water (19).
The aim of this study was the determination of hematological changes in common carp caused by thermal stress and the potential ability of adaptation with emphasis on the number of pseudoeosinophilic granulocytes.
The study was performed on 49 specimens of common carp (Cyprinus carpio Linnaeus, 1758), 22 males and 27 females, sampled in the Jablanica reservoir (Bosnia and Herzegovina) after floods in January, 2015. The reservoir is spread from the Konjic municipality to the municipality of Jablanica (43° 41’ 0″ N, 17° 51’ 0″ E) with an area of 13 km2 and 270 asl. Fishnets were used for fish sampling. Average length of all fish was 13.61 ± 1.34 cm and average body mass was 25.24 ± 6.96 g. All captured fish survived the transport to the laboratory. Fish were transported to the laboratory using barrels where water was continuously enriched with oxygen. The fish were randomly (with approximately equal gender balance) placed in two aquariums (control and experimental) with a total volume of 100 L, equipped with aerators (CHAMPION CX-0098) and heaters (PEAR AQUATICS Y 978). Adaptation lasted 20 days and during that period temperature was set at 10°C for both groups (Laboratory of Physiology, Faculty of Science, Sarajevo, Bosnia and Herzegovina). All fish were fed with Eco FeedEx C 48/10 (Eco Feed Ltd, Serbia). Water monitoring was done daily and included: changing the filters, analysis of oxygen concentration by Winkler’s method (20) and analysis of ammonia concentration by Nessler’s method (20). This research included 25 control and 24 experimental specimens.
Control specimens (11 males and 14 females) were kept all the time at 10°C, while the water in the aquarium with experimental specimens (11 males and 13 females) was gradually heated to 28°C and at this temperature fish were kept for 30 minutes (21). After that time, the sampling of blood was performed by cardiac puncture. Before puncture of the heart, the fish (both groups) were anaestethised by isoeugenol in concentration of 40 mg/L (22). Blood sampling was performed by heart puncture with a sharp and sterile needle (1.0 to 1.2 mm) under sterile conditions.
The values of basic morphometric features, total and standard length, as well as the mass were obtained. Technical scales were used for body mass estimation, and the standard and total length were measured by an ichthyometer.
The analysis of the hematological parameters was performed with native blood without anticoagulant and included the number of leukocytes (WBC) and the morphological differentiation of leukocytes in peripheral blood: segmented (Seg) and nonsegmented neutrophils (Nonseg), lymphocytes (Ly), monocytes (Mo) and pseudoeosinophils (Pse). The number of leukocytes was determined in a Neubauer chamber (hemocytometer) according to the method of Kekic and Ivanc (23) and morphological differentiation was performed using an Olympus BX41 microscope with Olympus DP12 camera.
Data are presented as means ± 1 SD accompanied by minimum, maximum and coefficient of variation (%). The Kolmogorov-Smirnov test was used to estimate the normality of data distribution. Student’s t-test was use to assess the differences between the groups. Statistical analysis were performed by SPSS (Version 20.0, SPSS, Inc., Chicago, IL, USA).
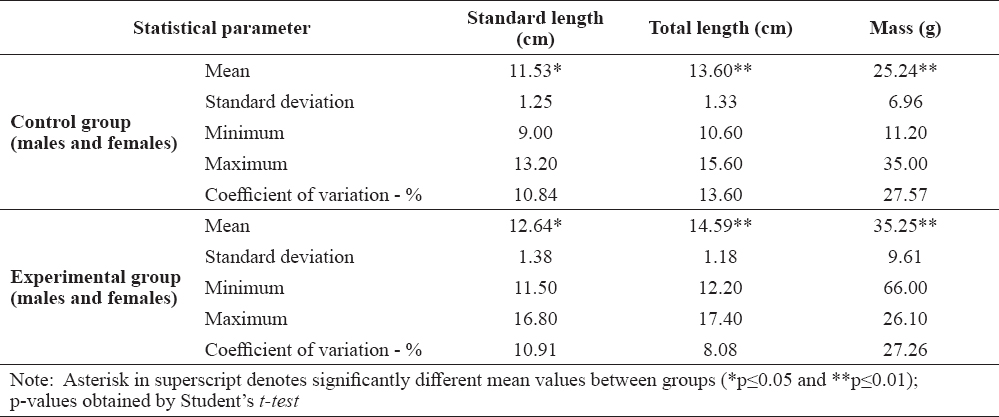
Descriptive statistics of the morphometric parameters and weight of control and experimental group is presented in Table 1.
Table 1. Standard length, total length and mass of control and experimental common carp
Kolmogorov-Smirnov test showed that all morphometric parameters had normal distribution (p>0.05).
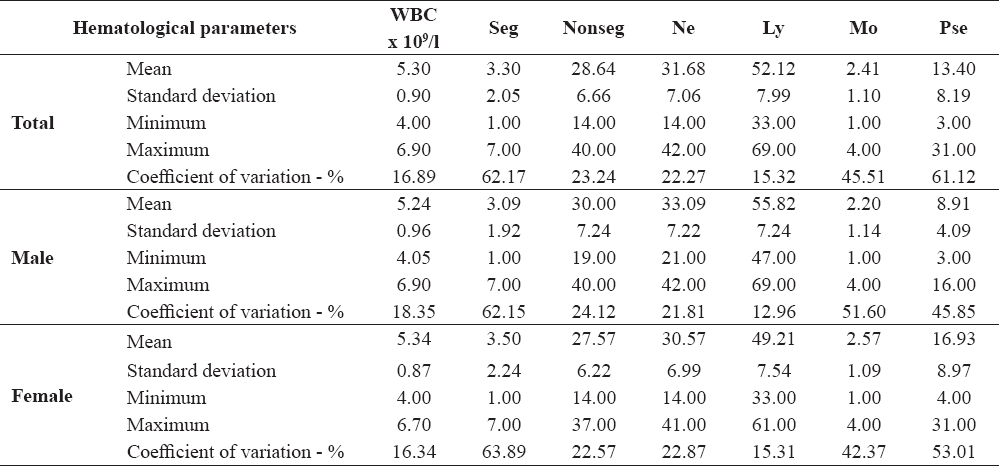
The analyzed white blood cells parameters of the control group are shown in Table 2.
Table 2. Hematological parameters of the control group of common carp
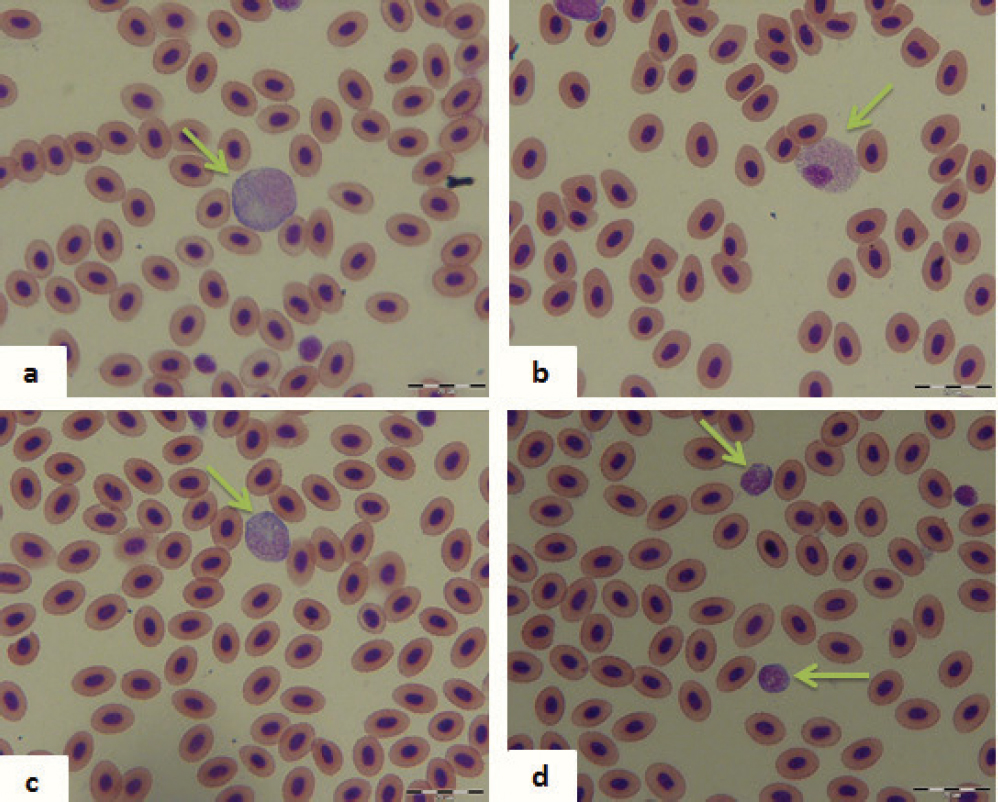
The analysis of the leukocyte number and differential blood count in the control fish showed high individual variation of Seg (CV=62.17%), Mon (CV=45.51%) and Pse (CV=61.12%). These variations were characteristic for both males and females. Females had a much higher number of Pse and less Ly compared to males. Number of Seg, Nonseg and Mon was approximately equal in control males and females. Types of leukocytes in common carp are shown in Fig. 1.
Figure 1. Types of leukocytes (green arrow) in common carp (Cyprinus carpio): a – monocyte; b – pseudoeosinophile; c – neutrophil; d – lymphocytes (magnification bar 20 μm)
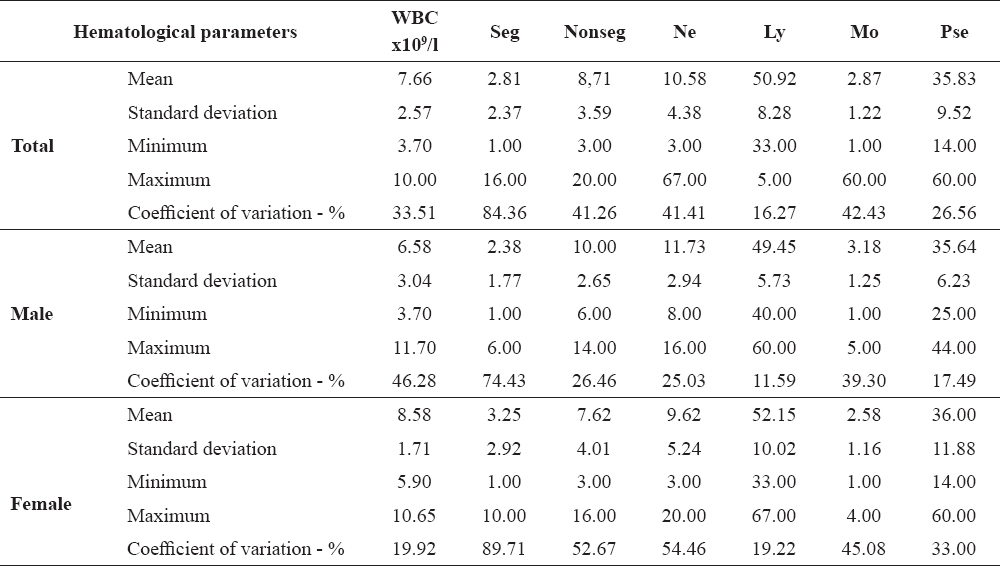
WBC parameters of the experimental group are shown in Table 3.
Table 3. Hematological parameters of the experimental group of common carp
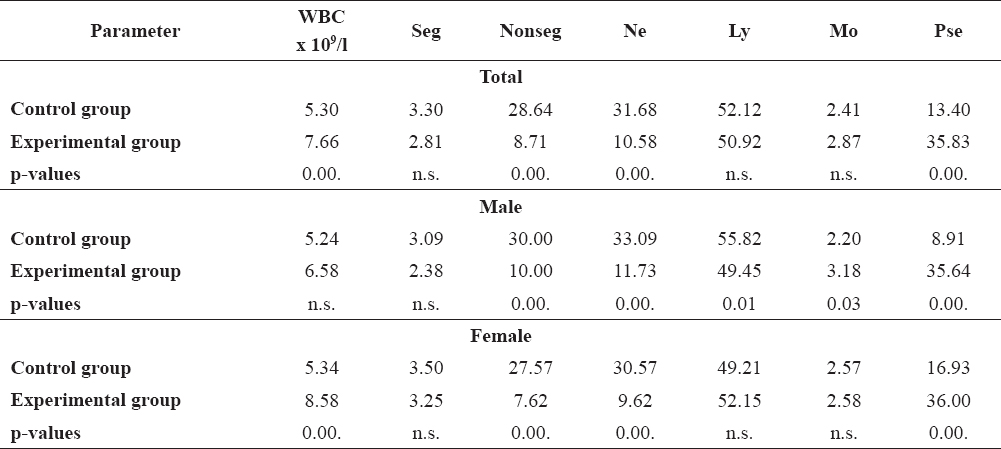
The experimental fish that were exposed to increased water temperature showed a wider distribution of the analyzed parameters in relation to the control group. This group was characterised by higher values of the coefficient of variation for Seg, Nonseg and Mon, except for the value of Ly and Pse. Larger variation of hematological parameters were noticed during the thermal stress. The largest variations were among the segmented neutrophils (CV=89.71% for females). The analyzed parameters of males showed moderate distribution in relation to females, except of WBC and Ly number. Gender specific analysis of mean values between control and experimental group is shown in Table 4.
Table 4. Gender specific analysis of mean values of common carp
Comparing the mean values of the control and experimental fish, there was an increase in the number of white blood cells, Mo and Pse, while Seg, Nonseg and Ly were decreased. Statistically significant differences (p=0.00.) were found for the WBC, Nonseg and Pse between control and experimental fish. Experimental group of males showed increased number of WBC, Mo and Pse, where significant differences were found for Nonseg, Ne, Pse (p=0.00.), Ly (p=0.01) and Mo (p=0.03). In experimental females, increase in total WBC number, Ly, Mo and Pse was evident, while significant differences (p=0.00.) were obtained for the WBC, Nonseg, Ne and Pse.
Adaptation mechanisms to water temperature are very important and they can be monitored by hematological analysis. To our knowledge, there is no available data for hematological parameters in similar conditions for the common carp, which makes our study even more important. The importance of this study is in the fact that Pse are one of the most present form of leukocytes in the common carp’s blood. It is considered that these types of cells can take the roles of other leukocytes and the function of cell is determined by a dominant type of granules (24). This research was confirmed by Hasković et al. (21). The hematopoiesis of that type of cell happens faster and only one type of cell proliferates. Our study showed that they were present in the blood and their number was increased during thermal stress, but they did not become the dominant form of leukocytes.
In the peripheral circulation of fish exposed to elevated water temperatures, an increased number of Pse can be observed as a result of catecholamines and cortisol effects, whose secretion was stimulated by thermal stress (25). Cortisol and catecholamines accelerate the maturation of granulocytes in hematopoietic centers, especially Pse, which were obviously the first hematological cells that enter into the circulatory system as a consequence of any stress. Interrenal chromaffin tissue produces a number of interleukins (IL3, IL3R, IL5) (25) colony stimulating factors and growth factors which stimulate the maturation and ejection of Pse in peripheral circulation. Previous studies on Tilapia and tench (22, 26) showed that Pse and Ly were dominant cells (more than 65%) in the blood which is consistent with our research.
In the case of monocytes, complex fish proinflammatory cytokines triggered mobilization (27, 28) and they cross from pronephros melanomacrophage centers to peripheral circulation (29). Decreased Ly count was probably result of the fact that these cells are involved in the cellular and humoral immune response and could not easily leave the anterior part of pronephros in which they were mainly localized, in relation to Pse and Mo. Ne count was also reduced, suggesting that this type of granulocytes, was not the first line of circulatory and hematological defense of common carp to thermal stress.
Total WBC of experimental males was very similar to WBC of control males, but the proportion of different leukocyte types was changed. This was not the case with the females, where the total number of leukocytes between the control and experimental group was significantly increased, accompanied also with a changed ratio of different types of leukocytes. Females were much more susceptible to thermal stress in comparison to males, because in addition to Pse increase, a significant increase (p<0.05) was observed for WBC, which was not the case in comparison to the gender-specific response to thermal stress in previous research on tench (21).
Based on the collected data, it is possible to draw the following conclusions: eosinophilic granulocytes, basophilic granulocytes and monocytes in common carp’s blood were sporadically present, which is consistent with literature data for tench (21). Short-term hyperthermia lead to an increase of leukocytes number, especially Pse (p<0.05) and Mo, while Ly, Seg and Nonseg (p>0.05) were reduced. Adaptation mechanism in common carp, caused by higher water temperature, manifests mostly with an increased number of Pse and a decreased number of Ne, though a more detailed explanation of this phenomenon we have not found in the literature. Hematological adaptation of fish to thermal stress is specific for each fish species and there is no uniform hematological response to thermal stress, even among cyprinids, because some cyprinid fish lack Pse in circulation, while the other have large number of Pse, basophils or eosinophils which sheds a completely different light on their thermal adaptation (17, 26).
1. Pickering AD, Endocrine-induced pathology in stressed salmonid fishFish Res 1993; 17: 35-50. http://dx.doi.org/10.1016/0165-7836(93)90005-R
2. Everly GS, Lating JM, A clinical guide to the treatment of the human stress response 2013; USA: Springer science & Business media; 250-286. http://dx.doi.org/10.1007/978-1-4614-5538-7
3. Iwama GK, Morgan JD, Barton B.A, Simple field methods for monitoring stress and general condition of fishAqua Res 1995; 26: 4273-282. http://dx.doi.org/10.1111/j.1365-2109.1995.tb00912.x
4. Kazlauskiene N, Vosyliene MZ, Physiological state of Atlantic salmon (Salmo salar Linnaeus 1758) and sea trout (Salmo trutta trutta Linnaeus 1758.)Acta Zool Lith 2004; 14: 448-51.
5. Mitrašinović M, Suljević D, Hematological status of chub fish Leuciscus cephalus (Linnaeus 1758.) from Krupica and Zeljeznica riversVeterinaria 2009; 58: 1-263-76.
6. Kroupova H, Machova J, Svobodova Z, Nitrite influence on fish: a reviewVet Med-Czech 2005; 50: 461-471.
7. Ritossa FM, A new puffing pattern induced by a temperature shock and DNP in DrosophilaExp Biol 1962; 18: 571-573. http://dx.doi.org/10.1007/bf02172188
8. Blank M, Khorkova O, Goodman R, Changes in polypeptide distribution stimulated by different levels of Em and thermal stressBiochem Bioenerg 1994; 17: 349-360. http://dx.doi.org/10.1016/0302-4598(87)80045-4
9. Samali A, Cotter TG, Heat shock proteins increase resistance to apoptosisExp Cell Res 1996; 223: 1163-170. http://dx.doi.org/10.1006/excr.1996.0070PMid:8635489
10. Samali A, Holmberg CI, Sistonen L, Orrenius S, Thermotolerance and cell death are distinct cellular responses to stress: dependence on heat shock proteinsFEBS Lett 1999; 461: 3306-310. http://dx.doi.org/10.1016/S0014-5793(99)01486-6
11. Samali A, Orrenius S, Heat shock proteins: regulators of stress response and apoptosisCell Stress Chap 1998; 3: 4228-236.PMCid: PMC312968
12. Ellis AE, Leukocytes in fishJ Fish Biol 1977; 11: 453-491. http://dx.doi.org/10.1111/j.1095-8649.1977.tb04140.x
13. Tavares-Dias M, Barcellos JFM, Peripheral blood cells of the armored catfish Hoplosternum littorale Hancock: a morphological and cytochemistry studyBraz J Morphol Sci 2005; 22: 215-220.
14. Tavares-Dias M, Moraes FR, Hematologia de peixes teleosteosRibeirão Preto 2004; 1: 144-
15. Lin S, Hematopoiesis L.I. Zon, Zebrafish hematopoietic developmentHematopoiesis: A Developmental Approach 2001; USA: Oxford University Press; 149-151.
16. Kapoor BG, Khanna B, Ichthyology handbook 2014; USA: Alpha Science International; 1-35.
17. Karalija E, Leukocyte formula of European chub (Leuciscus cephalus Linnaeus 1758) in seasonal aspect (Master thesis) 2007; Sarajevo (B&H): University of Sarajevo;
18. Velagić E, The effects of thermal stress on morphologic erithrocytes parameters and percentage presence of blood elements in Prussian carp (Carassius gibelio Bloch 1782)Master thesis 2013; Sarajevo (B&H): University of Sarajevo;
19. Suljević D, Mitrašinović M, Effects of thermal stress on glucose level in serum of carp and crucian carpVeterinaria 2009; 58: 3-4201-209.
20. Standard Methods for the Examination of Water and Wastewater 1975; 14th Edition. 410-Method 418A and 418B
21. Hasković E, Mehinović L, Suljević D, Hasković D, Hajdarević E, Glamuzina B, Differential blood count of tench (Tinca tinca Linnaeus 1758) in conditions of thermal stressVeterinaria 2013; 62: 3-4175-184.
22. Gladden JN, Brainard BM, Shelton JL, Camus AC, Divers SJ, Evaluation of isoeugenol for anesthesia in koi carp (Cyprinus carpio)Am J Vet Res 2010; 71: 8859-66.PMid:20673083 http://dx.doi.org/10.2460/ajvr.71.8.859
23. Kekic H, Ivanc A, A new direct method for counting fish blood cellsIchtiology 1982; 14: 155-58.
24. Đikić D, Lisičić D, Skaramuca D, Matić-Skoko S, Tutman P, Benković V, Horvat Knežević A, Gavrilović A, Skaramuca B, Blood cellular components in wild caught Muraena helena, L. 1758Cybium 2011; 35: 2149-156.
25. Harris J, Bird DJ, Modulation of the fish immune system by hormonesVet Immun Immunopath 2000; 77: 163-176. http://dx.doi.org/10.1016/S0165-2427(00)00235-X
26. Tavares-Dias M, A morphological and cytochemical study of erythrocytes, trombocytes and leucocytes on four freshwater teleostsJ Fish Biol 2006; 2: 155-157. http://dx.doi.org/10.1111/j.1095-8649.2006.01089.x
27. Reid SG, Vijayin MM, Perry SF, Modulation of catecholamine storage and release by the pituitary-interrenal axis in the rainbow trout (Oncorhynchus mykiss)J Comp Physiol 1996; 165: 665-676. http://dx.doi.org/10.1007/BF00301135
28. Cortes R, Teles M, Tridico R, Acerete L, Tort L, Effects of Cortisol Administered through Slow-Release Implants on Innate Immune Responses in Rainbow Trout (Oncorhynchus mykiss)Int J Genom 2013; doi:10.1155/2013/619714 http://dx.doi.org/10.1155/2013/619714
29. Engelsma MY, Hougee S, Nap D, Hofenk M, Rambout J, Van Muisvinkel W, Verburg-van Kemenade V, Multiple acute temperature stress affects leucocyte populations and antibody responses in common carp, Cyprinus carpio LFish Shellfish Immun 2003; 15: 397-410. http://dx.doi.org/10.1016/S1050-4648(03)00006-8
© 2016 Suljević D. This is an open-access article published under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 40, Issue 1, Pages 5-11, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.1515/macvetrev-2016-0091, 2017