Mac Vet Rev 2017; 40 (1): 13 - 20
 10.1515/macvetrev-2016-0092
10.1515/macvetrev-2016-0092
Received: 01 April 2016
Received in revised form: 15 August 2016
Accepted: 28 September 2016
Available Online First: 04 November 2016
Published on: 15 March 2017
Keywords: estrus responses, hormones, gilts, PGF2α
Estrus synchronization is a well-established tool for the reproductive management of a variety of livestock species. Manipulating the estrous cycle of domestic animals is thus essential for improving livestock productivity, as it helps to generate more income for the farmer through the utilization of assisted reproductive techniques like artificial insemination, in vitro fertilization and embryo transfer technologies. In gilts, estrus manipulation is being increasingly used globally for achieving programmed piglet production through batch farrowing, especially in today’s modern intensive piggeries (1).
In spite of this widespread use, currently available methods for swine estrus synchronization face a number of challenges which include depression in reproductive performance, low conception rates, increased stress due to handling, risk of occurrence of cystic ovaries (as in allyl trenbolone feeding), occurrence of silent heats, anovulatory estrus, high cost, scarcity of pharmacological agents used, and the requirement of excessive handling (2). Consequently, options for estrus synchronization in pig production are still very limited, in comparison to options available in the husbandry of other domestic animals, like cattle, sheep and goat.
In a number of domestic livestock species, prostaglandin F2α (PGF2α) treatment is capable of inducing regression of the corpus luteum (luteolysis) during the greater part of the luteal phase and is therefore employed for estrus synchronization and timed artificial insemination in cattle (3), sheep (4), and buffaloes (5). Contrastingly, in female swine a single treatment with PGF2α is not effective for inducing luteolysis until after Day 12 of the estrous cycle (6, 7), and therefore cannot be utilized for estrus synchronization. The reason for the species difference in luteolytic activity of exogenous PGF2αis still unclear (8).
Notable investigators who have studied the luteolytic effect of PGF2α in pigs include Diehl and Day (6). These authors were among the earliest researchers who reported that no luteolytic effect was observed when PGF2α was administered to cycling gilts on day 10 and 12 of the estrous cycle. These same authors also observed that PGF2α did show abortifacient effects when administered to pregnant gilts at 25 to 30 days post conception (6).
Subsequently, Hallford et al. (9) reported that PGF2α given on days 12 and 13 of the estrous cycle was luteolytic in gilts, while PGF2α administered on days 4 and 5 had minimal effects on the growth of the corpora lutea (CL). In furtherance to these works, Estill et al. (10) reported that although the pig corpora lutea had long been considered refractory to the luteolytic effect of PGF2α before day 12 of the estrous cycle, luteolysis may well be possible if repeated injections are given from day 5 through day 10.
In view of the foregoing, it is important to continually study the potentials of PGF2α as a possible agent for estrus synchronization in pigs by designing newer protocols until a very successful protocol is found. In addition to this, the need to look for viable alternative techniques for estrus synchronization in all domestic animals cannot be overemphasized particularly in today’s modern intensive global livestock industry.
The aim of this current study was therefore to investigate the possible outcome of single, double and triple intramuscular PGF2α treatments on estrus responses and hormonal profiles in crossbred (Large White x Landrace) gilts.
Sixteen crossbred (Large White x Landrace) pubertal gilts were used for this study. The animals were obtained from a reputable breeder in Ibadan, the capital of Oyo State, Nigeria. All the animals used for this study were between 7-9 months of age and were confirmed to be cycling via written records obtained from the breeder. The gilts weighed between 45 to 55 kg.
The gilts were housed in neat and well managed pens at the Teaching and Research Farm (TRF), University of Ibadan, Nigeria, and were fed on a standard corn/soy-bean/brewers’ dried grain diet, with clean fresh water supplied ad libitum, throughout the study period. Ethical approval for this work was obtained from the University of Ibadan Animal Care and Use Research Ethics Committee (ACUREC).
Four gilts each were assigned into Groups A, B, C, and D. Group A gilts were treated with Prostaglandin F2α (PGF2α) injection intramuscularly (I.M.) at a dose of 5mg per gilt; Group B gilts were treated with PGF2α I.M. at a dose of 7.5mg per gilt, and Group C gilts were treated with 10mg PGF2α I.M. per gilt; all on Days 0, 6 and 13 of the experiment.
Group D gilts formed the Control group and they were also handled similarly except that they were treated with I.M. injections of 2ml phosphate-buffered saline (PBS) instead of PGF2α.
Blood samples were collected for four days after each PGF2α injection, and heat detection via mounting trials was also conducted daily to detect estrus occurrence in test animals.
Blood was collected from the Vena Cava of the gilt using a 21-gauge needle into labeled (sterile) plain sample bottles and the blood was allowed to clot at room temperature by slanting the bottles. The serum was then harvested for hormone assay via centrifuging at 2000g for 15 minutes and decanting the supernatant into separate sterile sample containers. The resulting serum was then stored at -20°C until assayed for concentrations of progesterone, estradiol-17β, FSH and LH. Blood sampling was done for 4 days after each PGF2 treatment across all groups and the control.
All four hormones were assayed using commercially obtained ELISA kits (Pig LH, E2, Progesterone and FSH ELISA kits respectively) (Abnova Ltd, Taiwan) and assays were conducted according to manufacturer’s instruction. Assay sensitivity for LH assay was 0.08 ng/ml while assay sensitivity for estradiol was 5 pg/ml; with an intra-assay variation of 10.5% and an inter-assay variation of 8.5%. The sensitivity of the progesterone immunoassay was 1 ng/ml.
Mean (± S.E.M.) values of the hormones were calculated for all samples taken for the control and all PGF2α-treated animals. The collected data was then statistically assessed by analysis of variance (ANOVA) for two factorial, repeated measures. The Bonferroni test was applied for calculation of the statistical significance of mean differences. All statistical analysis was performed using GraphPad Prism version 5.01 (GraphPad Software, San Diego California USA, www.graphpad.com).
Results of the mounting trials conducted are summarized in Table 1. Of the twelve gilts treated with the different doses of PGF2α; four gilts (100%) from Group A came into estrus after the second PGF2α injection; while in Group B; two gilts (50%) exhibited estrus signs after the first injection, with the other two (50%) coming into estrus after the second injection. Only two gilts (50%) in Group C displayed signs of estrus during the study period and this was observed only after the second injection.
Table 1. Estrus responses of gilts in the various treatment groups to PGF2α injections
None of the gilts from all treated groups came into estrus after the third PGF2α injection. In addition, estrus signs were not shown by the control gilts throughout the study period.
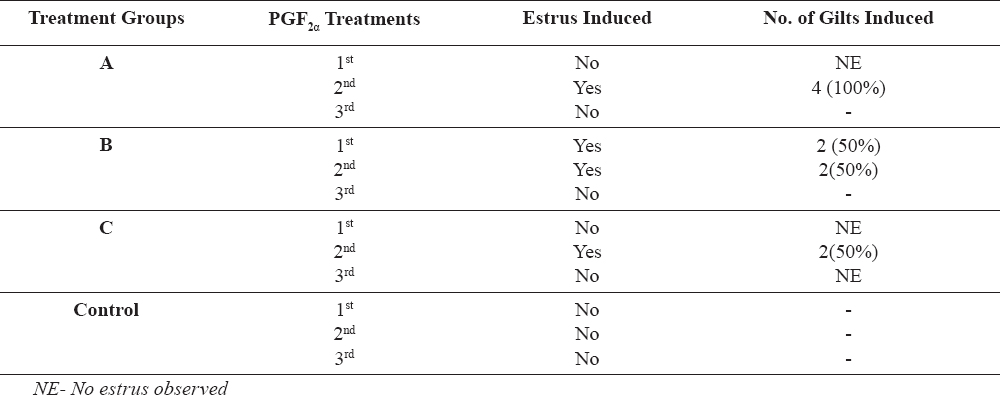
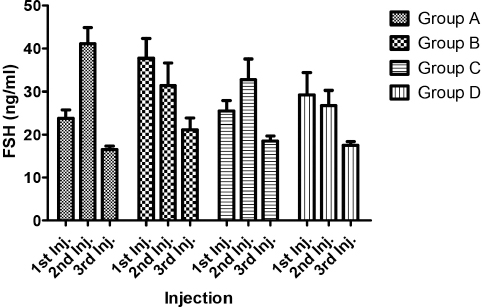
The mean concentration of all four hormones in sera was calculated for each day of blood sample collection. Figures 1-4 presents mean values of luteinizing hormone (LH), estradiol-17β progesterone (P4), and follicle stimulating hormone (FSH) respectively following the first, second and third PGF2α injections. The results showed that there was an increase in LH concentrations after the 2nd injection in all the PGF2α-treated groups (A,B,C), which tended to decline after the 3rd injection. Comparison of the LH results of PGF2α-treated groups with the control group showed that LH concentrations of Group A and Group B gilts differed significantly with the control; and these differences were observed after the 2nd PGF2α injection. LH values obtained from Group C gilts however did not differ significantly with the control across all 3 PGF2α injections.
Figure 1. Luteinizing hormone concentrations (ng/ml) of Test Gilts after 1st, 2nd and 3rd PGF2α injections
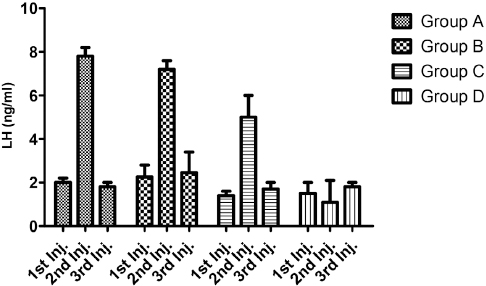
Figure 2. Estradiol-17β concentrations (pg/ml) of test gilts after 1st, 2nd and 3rd PGF2α injections
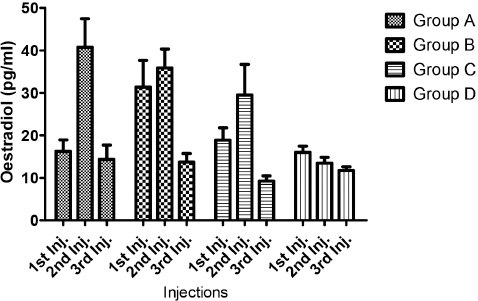
Figure 3. Progesterone concentrations (ng/ml) of test gilts after 1st, 2nd and 3rd PGF2α injections
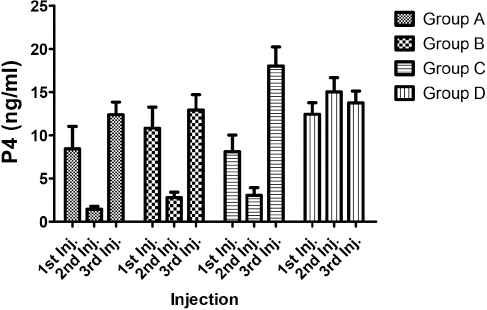
Figure 4. Follicle stimulating hormone (FSH) concentrations (ng/ml) of test gilts after 1st, 2nd and 3rd PGF2α injections
Estradiol values also showed a similar pattern of increased concentrations across all PGF2α-treated groups after the second injection with significant difference being observed after the 2nd PGF2αinjection in gilts of both Group A and Group B; when compared to the control group.
As per the results of the progesterone assay, there was a notable decrease in this hormone across all the PGF2α-treated groups corresponding with the days of serum collection after the second PGF2α injection; however these differences were not statistically significant when compared to the control gilts’ progesterone values.
The results of the FSH analysis showed that in Group A and C; there was an appreciable increase in the hormone after the second PGF2α injection which tended to decrease after the 3rd injection similar to what was observed in LH and estradiol. However, in Group B, mean FSH decreased gradually after each successive injection. Only FSH concentrations of Group A gilts were observed to be significantly different when compared with that of control gilts; and this difference was observed only after the second injection. There was no significant differences seen when the values for the two other groups were compared with the control.
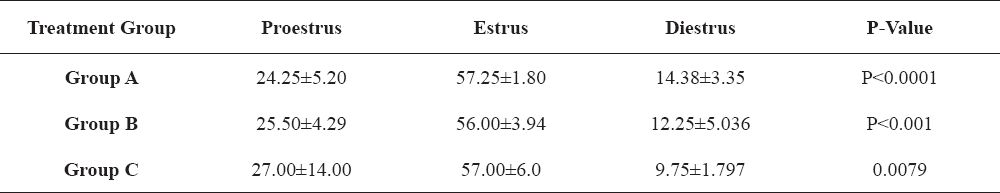
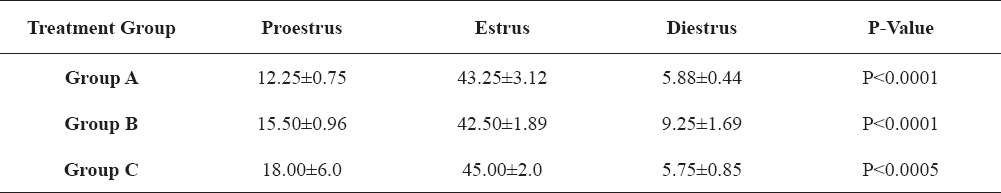
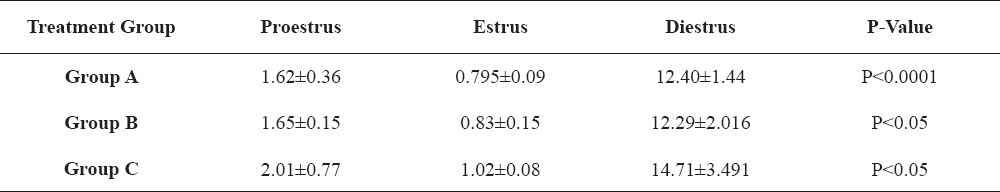
Table 2-5 show the changes in the four separate hormones assayed; LH, FSH, progesterone and estradiol, across the various stages of the estrous cycle in the PGF2α-treated animals that exhibited estrus. These results show that there was statistically significant differences (P<0.05) in the concentrations of the hormones across proestrus, estrus and diestrus periods of all the PGF2α-treated groups.
Table 2. Changes in luteinizing hormone concentrations (ng/ml) in PGF2α-synchronized gilts in relation to stage of estrous cycle
Table 3. Changes in follicle stimulating hormone (FSH) concentrations (ng/ml) in PGF2α-synchronized gilts in relation to stage of estrous cycle
Table 4. Changes in progesterone concentrations (ng/ml) in PGF2α-synchronized gilts in relation to stage of estrous cycle
Table 5. Changes in estradiol-17β concentrations (pg/ml) in PGF2α-synchronized gilts in relation to stage of estrous cycle
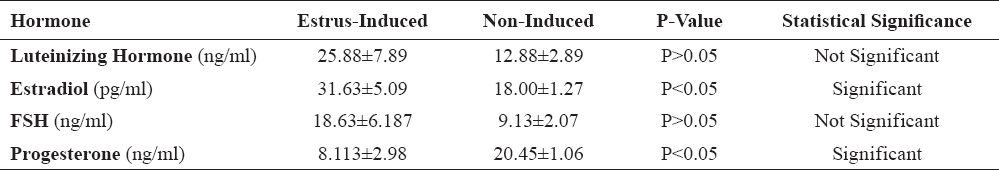
Table 6 compares the differences in hormonal profiles between estrus-induced and non-induced PGF2α-treated gilts in this study. In summary, there were significant differences (P<0.05) in the oestradiol-17β and progesterone concentrations, while luteinizing hormone and follicle stimulating hormone concentrations did not differ significantly.
Table 6. Differences in hormonal profiles of estrus-induced and non-induced PGF2α-treated gilts
This study has shown that it is possible to induce estrus in post-pubertal cycling gilts by administration of PGF2α intramuscularly seven days apart. The results of the mounting trials across the treatment groups showed that all four gilts (100%) in group A came into estrus at the same time; i.e. 4 days (96 hours) after the second injection, while in group B, two of the gilts (50%) came into estrus after the first injection; and the other two (50%) exhibited estrus following the 2nd injection. The result for group C further showed that only two gilts (50%) displayed estrus signs, and that occurred following the second injection of PGF2α. Control gilts on the other hand, did not express behavioral estrus during this study, suggesting that PGF2α may have induced estrus in the treated animals. Estrus responses of gilts to respective PGF2α injections in the present investigation showed that with the exception of two gilts in Group B; the PGF2α-treated animals generally did not exhibit estrus behavior after the first injection, but majority of them (66.6%) expressed signs of behavioral estrus after the second injection. This suggests that the second PGF2α injection was more effective than the first and third injections in causing estrus in the test gilts. However, it could also be that there was a cumulative effect of repeated administration of PGF2α which thus resulted in most of the gilts expressing estrus after the second injection. In addition, of all the three different doses used in this present investigation, it was demonstrated that the 5mg PGF2α dose was the most effective in terms of estrus synchronization because it was in this group that the highest proportion of gilts came into estrus at the same time. It is also evident from this result that administration of the three different dosages of PGF2α injected intramuscularly seven days apart was sufficient in bringing into estrus at least 50% of all the treated gilts in each group. However, the reason why the most successful synchronization was observed in the 5mg-treated group is still quite unclear. Since PGF2α is not a specifically recommended drug for estrus synchronization in pigs, there is currently no recommended dose for synchronization in this species. It is therefore speculated here that the effective dose for inducing luteolysis and estrus in this species is not likely to be far from the 5 mg per gilt mark as shown by the result of the present study. These findings however conflict with an earlier report by Diehl and Day (6) who reported that 2mg and 5mg dose levels of PGF2α was not luteolytic when administered during the luteal phase of the estrus cycle. With respect to this study, the reason for the disparity between the findings of the current study and the report of Diehl and Day (6) could have been because of the fact that the earlier authors administered PGF2α once; i.e. only on Day 4 or 10 of the estrus cycle of pigs while the present study administered PGF2α thrice, 7 days apart. The singular dosing regimen of Diehl and Day (6) was most likely responsible for the lack of luteolytic effect observed in their study. Furthermore, with regards to differences in estrus responses due to dose of PGF2α in treated gilts, Puglisi et al. (11) had earlier reported that PGF2α-induced luteolysis was not related to the dose used or to the weight of the animals, but was dependent upon the maturity of the animals; i.e. whether prepubertal or mature gilts. Their results showed significantly greater luteal regression was observed in pre-pubertal gilts than in mature gilts. In this present study, there was no comparison between pre-pubertal and post-pubertal gilts, so it was not possible to confirm this finding based on the investigation conducted.
The results of LH and estradiol-17β concentrations showed that there was a significant difference observed in the Group A and Group B gilts when compared to the control gilts following the 2nd PGF2α injection. In all the treated groups, it was also observed that serum concentrations of LH, FSH and estradiol-17β reached maximal levels just prior to expression of estrus. These findings agree with the documented hormonal profile of naturally cycling estrus gilts which is reported to be characterized by a rise in estradiol and LH surge immediately prior to estrus expression in most domestic animals including pigs (12, 13).
Progesterone concentration, though not significantly different across the groups after the respective PGF2α injections, showed a notable decrease in concentration across all the PGF2α-treated groups corresponding with the days of serum collection after the second PGF2α injection; which incidentally was the period when most of the test gilts in this study expressed estrus behavior. What this signifies is that during PGF2α-induced estrus in pigs, it is expected that progesterone, the major hormone of the luteal phase, will decrease appreciably. This finding is also corroborated by the works of Noguchi et al. (12) who reported also that plasma progesterone concentrations dropped to less than 1 ng/ml 30-48 hours in pseudopregnant swine after treatment with cloprostenol (a PGF2α analogue).
Changes in the concentrations of all the hormones in the animals that expressed behavioral estrus also showed that there was a significant difference in all the four hormones across all the PGF2α-treated groups. While there was a marked increase in LH, FSH and estradiol during proestrus and estrus; progesterone was highest during the diestrus period in all the estrus induced animals. These findings agree with the findings of Noguchi et al. (12) who previously used PGF2α to synchronize estrus in estradiol dipropionate (EDP) induced pseudopregnant sows.
From the results of this study it is hereby concluded as that it is possible to synchronize estrus in cycling post-pubertal gilts if PGF2α is administered repeatedly seven days apart. In addition, treatment with 5 mg PGF2α was the most effective dose of synchronizing gilts when three repeated intramuscular injections of PGF2α were administered to cycling gilts. It is also advanced that if three injections of PGF2α are administered to gilts 7 days apart, most gilts are likely to come to estrus after the second injection. Furthermore, hormonal profiles of gilts synchronized using 3 injections of PGF2α are similar to hormonal profiles of naturally cycling gilts. Reproductive hormone sampling therefore is a good indicator of induced estrus in PGF2α-treated gilts.
Based on the result of this present study, it is hereby recommended that future studies should investigate the fertility, conception, birth weight and litter size of gilts synchronized with intramuscular PGF2α administered 7 days apart, as well as duration of PGF2α-induced estrus and optimum time for artificial insemination
1. Ziecik AJ, Biallowicz M, Kaczmarek M, Demianowicz W, Rioperez J, Wasielak M, Bogacki M, Influence of estrus synchronization of prepubertal gilts on embryo qualityJ. Reprod. Dev 2005; 51: 3379-384. http://dx.doi.org/10.1262/jrd.17008 PMid:15827382
2. Kirkwood RN, Pharmacological intervention in swine reproductionSwine Health Prod 1999; 7: 129-35.
3. De Rensis F, Peter A, The control of follicular dynamics by PGF2α GnRH, hCG and estrus synchronization in cattleReproduction in Domestic Animals 1999; 34: 49-59.
http://dx.doi.org/10.1111/j.1439-0531.1999.tb01383.x
4. Menchaca A, Miller V, Gil J, Pinczak A, Laca M, Rubianes E, Prostaglandin F2α treatment is associated with timed artificial insemination in ewesReproduction in Domestic Animals 2004; 39: 352-355. http://dx.doi.org/10.1111/j.1439-0531.2004.00527.x PMid:15367269
5. De Rensis F, Lopez-Gatius F, Protocols for synchronizing estrus and ovulation in buffalo (Bubalus bubalis): A reviewTheriogenology 2007; 67: 209-216.
http://dx.doi.org/10.1016/j.theriogenology.2006.09.039 PMid:17112577
6. Diehl JR, Day BN, Effect of prostaglandin F2α on luteal functiion in swineJ Anim Sci 1974; 39: 392-396. http://dx.doi.org/10.2527/jas1974.392392x PMid:4845714
7. Guthrie HD, Polge C, Luteal function and oestrus in gilts treated with a synthetic analogue of prostaglandin F2α (ICI79.939) at various times during the oestrous cycleJournal of Reproduction and Fertility 1976; 48: 423-425. http://dx.doi.org/10.1530/jrf.0.0480423 PMid:1033285
8. Manzoni C, Saleri R, Kirkwood RN, Macchi E, Mongkol T, De Rensis F, Double cloprostenol administration during mid-luteal phase of oestrous cycle does not modify the interoestrous interval in giltsItal J Anim Sci 2014; 13: 527-529.
9. Halfordm DM, Wettemannm RP, Turmanm EJ, Omtvedt IT, Luteal function in gilts after prostaglandin F2αJ Anim Sci 1975; 41: 1706-1710. http://dx.doi.org/10.2527/jas1975.4161706x
10. Estill CT, Britt JH, Gadsby JE, Repeated administration of prostaglandin F2α during the early luteal phase causes premature luteolysis in the pigBiol. Reprod 1993; 49: 181-185. http://dx.doi.org/10.1095/biolreprod49.1.181 PMid:8353186
11. Puglisi TA, Rampacek GB, Kraeling RR, Kiser TE, Corpus luteum susceptibility to prostaglandin F2α (PGF α) luteolysis in hysterectomized prepuberal and mature giltsProstaglandins 1979; 18: 2257-264. http://dx.doi.org/10.1016/0090-6980(79)90111-4
12. Noguchi M, Kashiwa S, Itoh S, Okumura H, Kure K, Suzuki C, Yoshioka K, Reproductive hormone profiles in sows on estrus synchronization using estradiol dipropionate and prostaglandin F2α-analogue and the reproductive performance in female pigs on commercial farmsJ. Vet. Med. Sci 2013; 75: 3343-348.
http://dx.doi.org/10.1292/jvms.12-0022PMid:23131781
13. Soede NM, Langendijk P, Kemp B, Reproductive cycles in pigsAnim. Reprod. Sci 2011; 124: 251-258. http://dx.doi.org/10.1016/j.anireprosci.2011.02.025 PMid:21397415
© 2016 Alimi Y.A. This is an open-access article publishedunder the terms of the Creative Commons Attribution License whichpermits unrestricted use, distribution, and reproduction in any medium,provided the original author and source are credited
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 40, Issue 1, Pages 13-20, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.1515/macvetrev-2016-0092, 2017