Mac Vet Rev 2018; 41 (1): 5 - 19
 10.1515/macvetrev-2017-0027
10.1515/macvetrev-2017-0027
Received: 02 June 2017
Received in revised form: 17 July 2017
Accepted: 30 August 2017
Available Online First: 19 November 2017
Published on: 15 March 2018
Keywords: genistein, pituitary gland, hormone producing cells
Phytoestrogens are considered to be plant-based compounds that elicit similar effects to estrogen, due to their structural and/or functional similarity. One major class of phytoestrogens are the lignans, which are components of plant cell walls. Other groups of phytoestrogens are phenolic compounds, of which the isoflavones and coumestans are the most researched groups (1). Even though phytoestrogens are abundant in huge number of plants, soybeans and soy based foods are the greatest source of isoflavones in human and animal diets. In diets, most of isoflavones are in a glycoside form that is biologically less active (genistin, daidzin) and these molecules are cleaved by intestinal bacteria to the aglycone forms (genistein, daidzein), which are then biologically active (2).
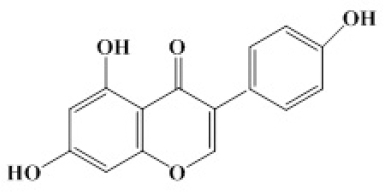
Genistein (G; Fig.1.) is one of the best characterized isoflavones. It was first isolated from soybeans in 1931 (3), while its estrogenic activity wasn’t identified until the 1950s (4, 5).
Figure 1. The chemical structure of genistein
Thus, it was found that G increased uterus weight in several animal species (5). G possesses two phenolic rings (B ring is linked to the heterocyclic ring at the C3) similar to estrogens, so it can act through an estrogen receptor (ER)-mediated mechanism, by increasing or decreasing the transcription of ER-dependent target genes (estrogenic, anti-estrogenic activity). It is able to bind to both type of ER (ERα and ERβ), however a higher potency toward ERβ is observed (6-8). Also, there is a difference in G bounding affinity for ER between in vivo and in vitro conditions. In vivostudy showed the equal bounding affinity of isoflavones to ERα and ERβ, while in vitro exhibited a higher binding affinity for ERβ (9). The effect of G is dose dependent, and whether they will act as estrogen agonists or as antagonists depend on the quantity of isoflavones (10). Precisely, isoflavones are associated with the ERs who are available (at a low level of endogenous estrogen), and the estrogen effect is potentiated with an injection of lower doses of isoflavones (11). Furthermore, it is important to emphasize that the estrogenic potency of G is much weaker than that of physiological steroids, actually only ~1/10,000 to 1/50,000 than that of estradiol (12). Genistein is a potent inhibitor of tyrosine protein kinases (PTKs), too (13, 14). Considering that PTKs inter alia catalyze phosphorylation of growth factors involved in tumor cell proliferation, genistein can potentially slow tumerogenesis. Inhibiting DNA topoisomerases, it can affect DNA replication (15, 16). Also, there is the ability of soy isoflavones to regulate androgen biosynthesis in Leydig cells due in part to action on Leydig cell 3β- hydroxysteroid dehydrogenase activity (17). Plus, isoflavones indirectly exert their antioxidant effect by activating enzymes antioxidant protection: catalase, superoxide dismutase and glutathione peroxidase (18). Genistein is able to bind to androgen receptor (AR), as well. However, its AR-binding affinity is significantly lower as compared with testosterone and estradiol (19).
Isoflavones are recommended to have cancer-protective properties. Consequently, women whose diet is isoflavone-rich have a reduced risk and rates of breast cancer (20, 21). Also, the significant inverse association between breast cancer risk and urinary excretion of the isoflavone metabolite equol were observed (22). Shimizu et al. (23) study indicates the rate of breast cancer in immigrants from Japan after years of living in North America, as well as their daughters is almost the same as in the indigenous Americans, which highlights the importance of diet in the etiology of cancers. Besides the genistein-provoked inhibition of breast cancer cells proliferation, a similar effect was identified in prostate and colon cancer (24). Studies carried out on people of Japanese origin living in Hawaii, pointed the reduced risk of prostate cancer as a result of a diet rich in soy products (25).
Numerous data support the hypothesis that genistein has an impact on reducing the risk of cardiovascular disease, as a cardio-protective compound (26). The reduced level of lipids (30%-40% lower level of low-density lipoprotein-LDL) provoked by isoflavones, is at least in part responsible for the cardio-protective effect (27). As we have mentioned above, G is a powerful antioxidant too, and it is able to inhibit the oxidation of LDL, in vitro. Previous data showed that isoflavones prevent the formation of plaques in blood vessels of patients with atherosclerosis and inhibit the adhesion of cells, and in this way act as an antiatherogenic factor (28). The results of our research group indicated lower cholesterol levels and increased level of triglycerides in male rats after treatment with genistein and daidzein, which can reduce enthusiasm for their application (29).
Some studies suggest a possible role of soy isoflavones in bone metabolism and osteoporosis prevention (30, 31). Namely, in postmenopausal women who have consumed the soy enriched food the increase in bone mineral content has been observed. (32). Also, by reducing the level of alkaline phosphatase in the serum, genistein exerts its osteoprotective effect (33).
Similarly, we should not ignore the beneficial effect of isoflavones on menopause symptoms, such as heat waves (“hot flushes”), loss of vaginal secretions, poor sleep and anxiety accompanied by increased secretion of stress hormones (34). Our research has detected a lower level of stress hormone in old rats treated with G, which increases the interest in genistein application in andropause (35). Also, soy isoflavone prevented the development of adenocarcinomas in the prostate and seminal vesicles in a rat carcinogenesis model. Plus, the soy diet reduced the growth of transplantable prostate adenocarcinomas and inhibited tumor cell proliferation and angiogenesis of transplantable prostate cancer (36).
The estrogenic action of G has been examined and confirmed in two major estrogen responsive organs, the uterus and the ovaries (37, 38). More precisely, neonatal exposure to genistein caused a dramatic increase in ERα in granulosa cells in mice (37). The results from our study showed that ovaries exposed to genistein and daidzein have had healthier primordial and primary follicles and less atretic follicles, which means they increased the ovarian follicular reserve by stepping down transition from primordial to primary follicles and less atretic follicles in middle-aged rats (39). Also, we observed that genistein and daidzein exposure have decreased superoxide dismutase and catalase, indicating that soy phytoestrogens acted as free radical scavengers, which highlight their antioxidant potential (39). Increased serum progesterone level was observed following genistein and daidzein treatment, as well (39).
In addition to the mentioned benefits of isoflavones use, it has been understood for a long time that consumption of hormonally active plant constituents can adversely affect reproduction in domestic animals, up to induction of permanent infertility (40). Considering that estrogen can stimulate breast cancer cells, there are concerns that isoflavones may not be safe for women who have or already had breast cancer. While isoflavones in general should have an anti-estrogenic effect by blocking real estrogen, some studies in animals have found evidence that, under certain circumstances, soy isoflavones might stimulate breast cancer cells (41, 42). Also, Unfer at al. (43) showed that long-term treatment with soy phytoestrogens increased the occurrence of endometrial hyperplasia, which calls into question the long-term safety of phytoestrogens with regard to the endometrium. Human pre-natal exposure to isoflavones may affect human semen quality produce or contribute to male infertility, cause birth defects in male infants, breast and testicular cancers, obesity and other adverse health effects (44). Goldin et al., (45) presented that isoflavones might slightly decrease testosterone levels in man, as well. Further concerns relate to soy’s potential effects on the thyroid gland. Thus, in patients with impaired thyroid function soy products have been observed to reduce absorption of thyroid medication (46). Also, in people who are iodine deficient isoflavones may directly inhibit the function of the thyroid gland (47). Soy isoflavones’s consummation had no effect on thyroid hormone levels or actually increased levels in humans and animals, which make things more confusing (48). Considering that isoflavones are able to derail the normal endocrine signaling, they are classified into the category of an endocrine disruptor (49).
The central neuroendocrine systems are responsible for the control of homeostatic processes, including reproduction, growth, metabolism and energy balance, as well as stress responsiveness. The pituitary gland, as part of the neuroendocrine system, is often dubbed the “master gland” because its hormones control other parts of the endocrine system, namely the thyroid gland, adrenal glands, ovaries, and testes (50).
Anatomically, the pituitary is divided into adenohypohysis and neurohypophysis. The adenohypohysis is represented by the anterior lobe, the intermediary lobe and the infundibulo-tuberal lobe in mammals. Rat anterior pituitary lobe contains five types of hormone-producing (HP) cells: somatotrops (growth hormone - GH cells), lactotrophs (prolactin - PRL cells), corticotrops (adrenocorticotropin hormone - ACTH cells), thyrotrophs (thyroid stimulating hormone - TSH cells) and gonadotrophs (follicle-stimulating hormone - FSH and luteinizing hormone - LH cells, 51). Pituitary hormone synthesis and secretion represent the integrated processes, controlled by hypothalamic, intrapituitary and peripheral signals. Thus, GH secretion from pituitary somatotrophs is controlled by the interplay of growth hormone releasing hormone (GHRH) and somatostatin (SS), secreted from the hypothalamus (52). Prolactin, the pituitary hormone whose principal control is inhibitory, is regulated by dopamine (causing inhibition of prolactin secretion) and TRH (stimulated prolactin secretion) (51). The corticotrophin-releasing hormone (CRH) and arginin-vasopresin from hypothalamus stimulate the release of ACTH from the pituitary, which in turn acts on the adrenal cortex to trigger the release of glucocorticoids (53). Thyreotropin releasing hormone (TRH) from the hypothalamus stimulates TSH cells to secrete TSH, which in turn stimulates the thyroid gland to produce thyroxin. The hypothalamus through the secretion of gonadotrophin-releasing hormone (GnRH) governs the activity of the pituitary gland, an organ which serves as an amplifier, transmitting the brain signal via the secretion of LH and FSH, to act on the gonads (54).
Morphology of rat HP cells: GH cells are ovoid, sphericalor polyhedral cells with an eccentric nucleus and abundant, densely granulated cytoplasm. PRL cells are large, ovoid angular cells with an eccentric nucleus, more numerous in the female animals. ACTH cells are angular cells, sending cytoplasmic processes among neighboring cells, while nucleus follows the shape of the cell body. TSH cells are angular cells, too. Gonadotrophs are immunoreactive for both FSH and LH or only for one of these hormones. In females half of these cells are bihormonal, while in males almost all cells contain both hormones (55-59).
Dada et al. (56) analyzed cell types in pars distalis of the normal young rat with respect to their percentages and the relative volumes they occupy. In rats the percentages of cell types were: PRL approximately 50 %, GH 20 %, LH 5%, FSH 4%, ACTH 3% and TSH 2%. However, this proportion is not represented in all studies dealing with percentage of hormone producing cells, due to different methods used for analyses, different sex, age, and species. The proportion of HP cells type changes dynamically, as a response to different endocrine demands (60, 61). Previously, it was thought that a particular type of HP cells synthesizes only one hormone and according to this principle cells are characterized as somatotrops, lactotrophs, etc. However, when the secretory granules of a certain HP cells revealed several hormones, the concept of the plurihormonalni HP cells was accepted. It was shown that 30% of all adenohypophysis HP cells in rats, mice and monkeys express mRNA for a variety of hormones (62). For example, subpopulation of gonadotropes and lactotropes also express GH (63).
Most of pituitary HP cells are targeted by estrogens, through direct and/or indirect mechanisms. The direct action of estrogen is enabled due to the expression of ER in almost all type of pituitary cells (64). Thus, Yin et al. (65) and Milosevic et al. (66) observed that estradiol provoked: PRL cell proliferation by direct action and LH secretion by positive feedback action (indirect action) in female rats. Results from our study indicated the decreased relative intensity of fluorescence and smaller volume of middle-age female rat FSH and LH cells, while PRL cells were more numerous, following estradiol treatment (0.625 mg/kg b.w.; 39). Also, stereological analysis has shown higher values of total number of rat ACTH cells, which was accompanied with the higher ACTH level, indicating the elevated synthesis and secretion of rat ACTH after estradiol treatment (67). On the other hand, the volume density of rat GH cells was reduced; total number of cells was unchanged, while the relative intensity of fluorescence of GH cells and GH blood level increased after estradiol treatment (0.625 mg/kg b.w.; 68). Previous work from our laboratory indicated inhibition of TSH cells in middle-aged female rats after chronic application of estradiol (0.625 mg/kg b.w; 69). From the all above it can be unambiguously concluded that estrogen impacts, directly or/and indirectly not only the function of HP cells, but also their stereological characteristics.
Due to structural and functional resemblance to the estrogen, the question may raise of whether and how G affects the morpho-functional features of pituitary HP cells. Considering that cell counts (stereology approach) have been essential in the discovery of important concepts in a variety of cell biology investigation, these findings are irreplaceable for the complete explanation of the impact of genistein on pituitary HP cells (70).
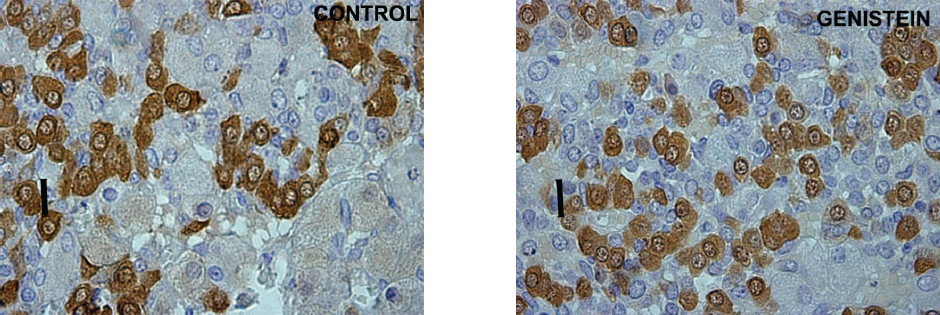
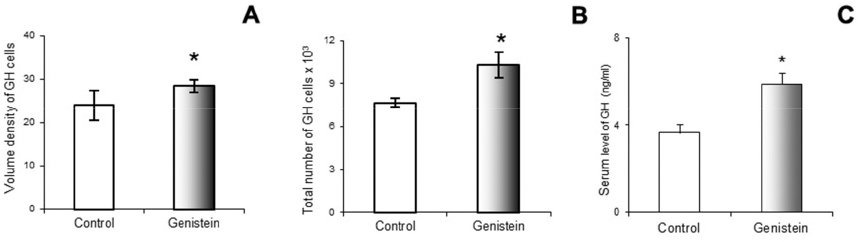
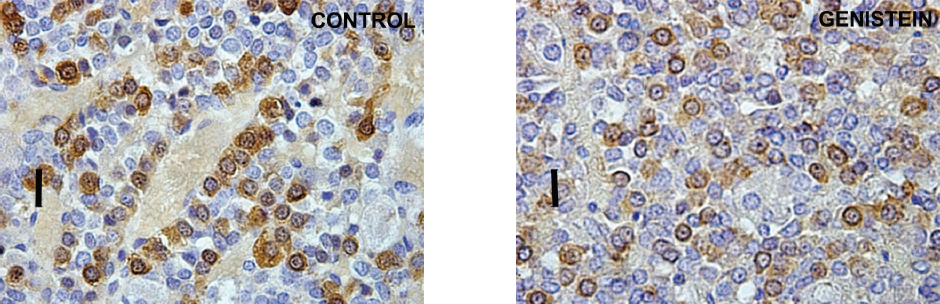
Using a stereological system (newCAST) Trifunovic et al. (68) showed the increase of pituitary volume following chronic G treatment (30 mg/kg b.w.) in adult orchidectomized rats. Orchidectomized adult animals were used experimentally to minimize steroids influence and separate the effect of sex steroid and genistein. The same study has indicated the greater volume density and total number of pituitary GH cells, as well as GH blood level following G treatment. The increases of stereological parameters of GH cells are a result of G indirect action via GHRH and direct via ER, expressed in GH cells (71; Fig. 2 and Fig. 3).
Figure 2. Immunoreactive somatotropes in the pituitary gland in control and genistein treated adult male rats. Scale bar 16 μm
Figure 3. Volume density of somatotropes (GH cells; A); Total number of GH cells (B); Blood GH concentration (C) in control and genistein treated adult male rats. The values are means ± standard deviation, n = 6 animals per group; * p<0.05 vs. control
More precisely, genistein action led to a higher volume density of GHRH in the Arc nucleus (72), probably operating through ER-dependent mechanisms, considering that 70% of those neurones express ER (73). If we compare the action of estradiol and genistein on morphfunctional parameters of GH cells, we can conclude that both treatments follow a similar course, the increase of GH cells action.
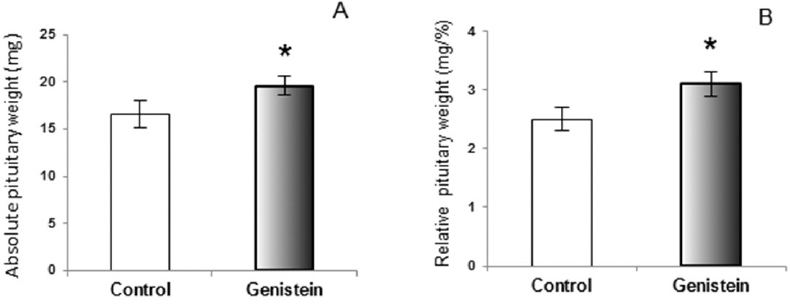
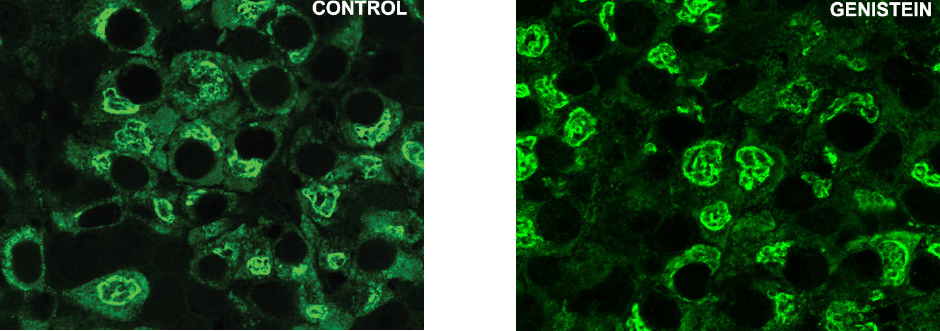
The similar conclusion, that genistein was an effective stimulator of GH secretion, was reached in the study on ewes (74). In orchidectomized middle-aged rats (animal model of andropause) genistein application provoked the increases of absolute and relative pituitary weight, which is explicable by their presumable estrogenic effect on the pituitary prolactin cells (75). Weaker immuno-histochemical staining and immuno–fluorescent signal have been observed within GH cells in middle-age rats following G treatment, which is a different effect compared to adult rats (Fig. 4 and Fig. 5).
Figure 4. Absolute pituitary weight (A); Relative pituitary weight (B) in control and genistein treated andropausal rats. The values are means ± standard deviation, n = 6 animals per group; * p<0.05 vs. control
Figure 5. Immunoreactive somatotropes in the pituitary gland in control and genistein treated middle-aged rats. Scale bar 16 μm
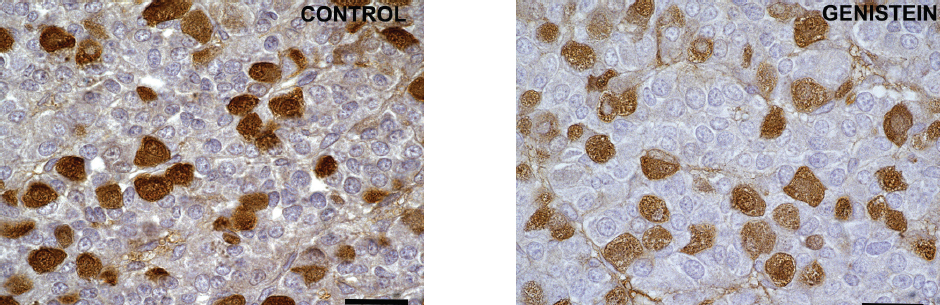
The Romanowicz et al. (76) data demonstrated that genistein may effectively modulate LH and PRL secretion in ovariectomized ewes, too. The data obtained in our experimental study indicated that genistein didn’t change relative intensity of the fluorescent signal, while alterations in cell morphology were observed in gonadotropic and lactotropic cells in middle aged female rats (Fig. 6). In fact, gonadotropic and lactotropic pituitary cells were changed in shape, with unevenly stained cytoplasms which gave cells the appearance of vacuolization (39).
Figure 6. Immunoreactive lactotrops in the pituitary gland in control and genistein treated middle aged female rats. Scale bar 50 μm
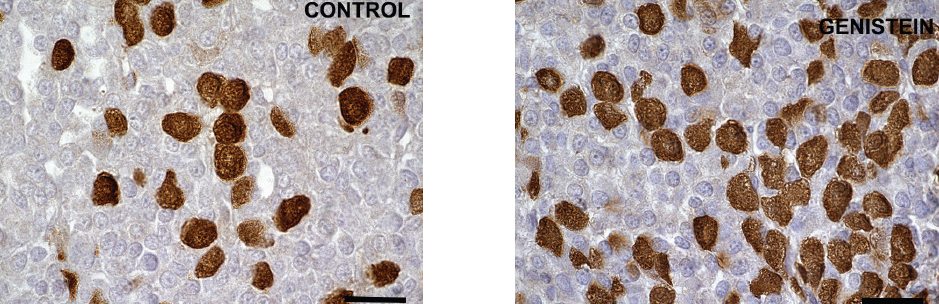
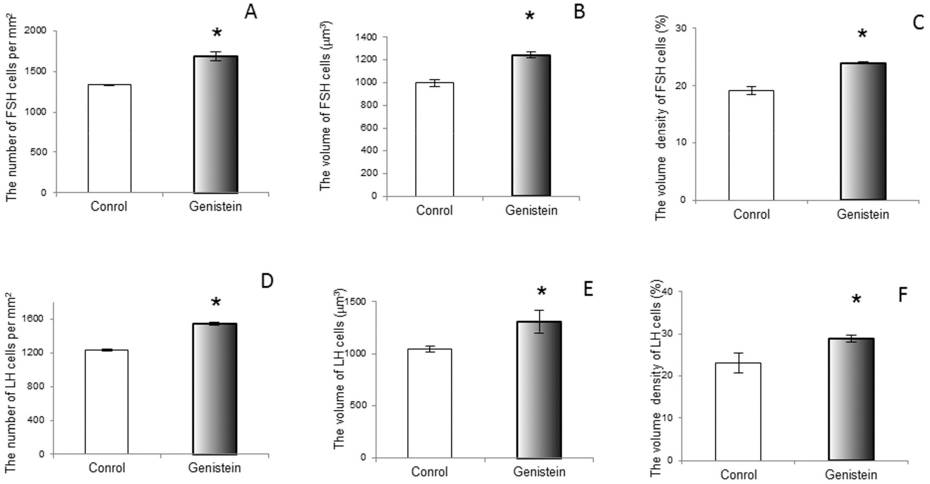
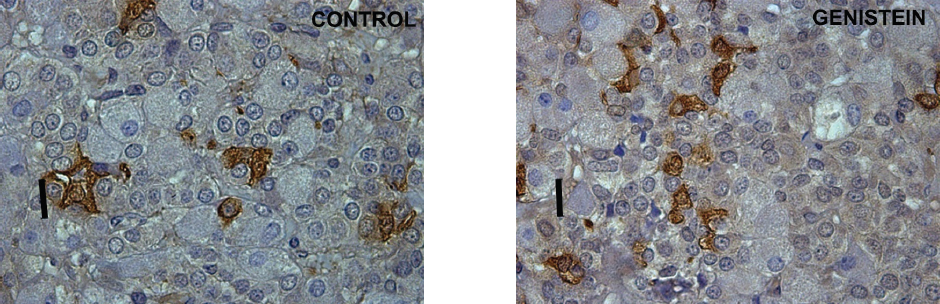
Considering the presence of various forms of ERs (with the serious potential of phytoestrogen sensitivity) in the pituitary gonadotropic and lactotropic cells, not only in the nucleus and cytoplasm but also in the membrane of rough endoplasmic reticulum and secretory vesicles, while keeping in mind that vesicular trafficking crucially defines cell mechanical status (77, 78), we can assume that the observed changes in cell morphology may be mediated by both genomic and non-genomic pathways. Short term exposure of immature female rats to genistein (50 mg/kg/ b. w.) induced significant increases in the number of FSH cells (by 21%) and LH cells (by 20%; 79). The study showed the increased volumes of FSH and LH cells by 19% and 20% and their volume densities by 20% and 20%, so in this experimental model genistein acted as an estrogen agonist (Fig. 7; Fig. 8 and Fig. 9).
Figure 7. Immunoreactive follicle-stimulating hormone producing cells in the pituitary gland in control and genistein treated immature female rats. Scale bar 20μm
Figure 8. Immunoreactive luteinizing hormone producing cells in the pituitary gland in control and genistein treated immature female rats. Scale bar 20μm
Figure 9. The number of follicle-stimulating hormone producing (FSH; A) and luteinizing hormone producing (LH; D) cell per mm2 (A); The volume of FSH (B) and LH (E) cell; The volume density of FSH (C) and LH (F) cell in control and genistein treated immature female rats. The values are means ± standard deviation, n = 6 animals per group; * p<0.05 vs. control
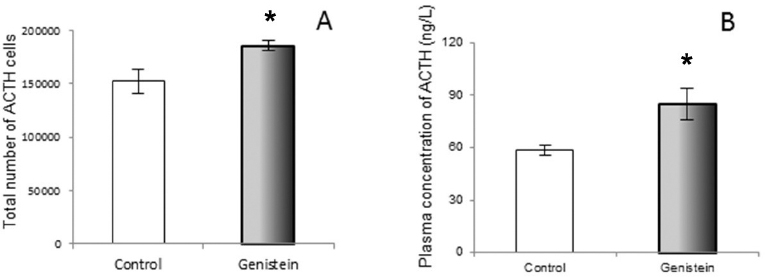
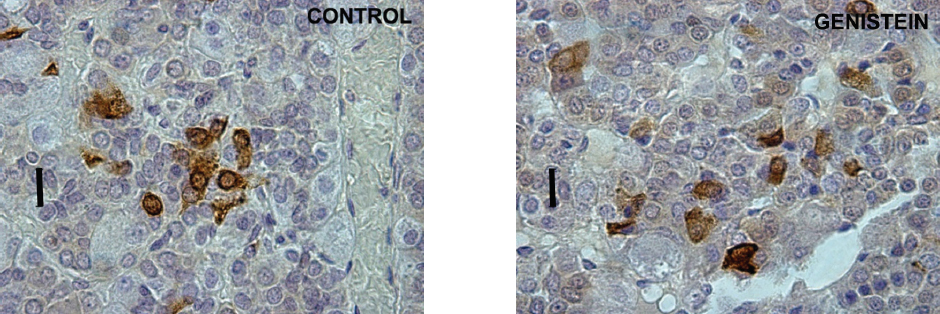
The serum concentration of FSH remained unchanged, while LH concentration increased after long-term administration of G to immature male rats (80). The previous study confirmed the enhanced LHβ mRNA expression and a greater proportion of LH cells after intracerebroventricular administration of G in ewes (81). However, in intact anestrus ewes LH cell percentage fell, but the associated increase of LHβ mRNA was presumed to be due to the stimulated rapid release of the hormone, following G exposure (82). In aromatase knockout mice, oral or subcutaneous, genistein exposure reduced serum FSH and LH concentrations i.e., back to the levels found in wild type mice (83). When it comes to ACTH cells of adult rats, the chronic G treatment caused an increase in the total number of ACTH cells (67). The plurihormonal cells concept is probably responsible for the increased ACTH cell number. Namely, plurihormonal cells which contain both ACTH and gonadotropic hormones were detected within the population of rat pituitary corticotrophs (84). The unchanged volume of particular ACTH cells after chronic genistein application, together with reduced immunostaining intensity and elevated circulating ACTH level, most probably represent the consequence of the continuous synthetic and secretory activity of these cells, due to constant hypothalamic stimulation (Fig. 10 and Fig. 11).
Figure 10. Immunoreactive corticotropes in the pituitary gland in control (C) and genistein treated (G) adult male rats. Scale bar 16 μm
Figure 11. The total number of corticotropes (ACTH cells; A) and plasma concentration of ACTH (B) in control and genistein treated adult male rats. The values are means ± standard deviation, n = 6 animals per group; * p<0.05 vs. control
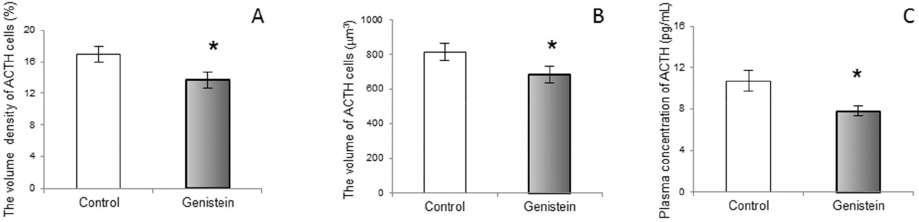
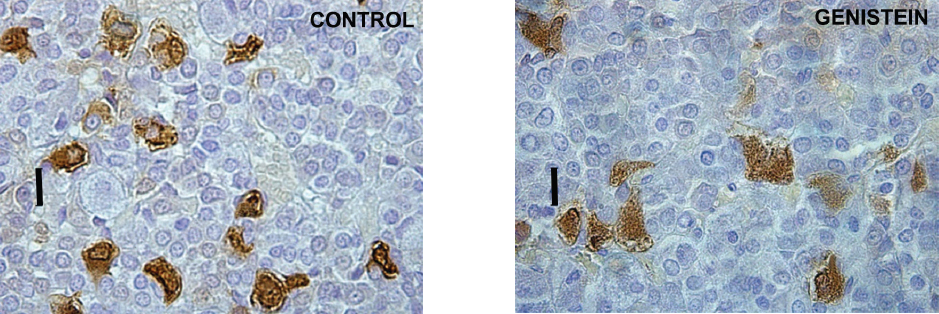
In contrast with the cited results, our work in middle-aged rats has shown the suppressive effects of genistein exposure on morphological parameters of ACTH cells, together with a lowering of blood ACTH levels (85; Fig. 12 and Fig. 13). Also, in vitro studies have shown the inhibitory action of genistein on rat pituitary cell proliferation (86). A likely explanation is the age-dependent decline in pituitary responsiveness to CRH, i.e. in middle-aged rats pituitary responsiveness is about 60% of that observed in young populations (87).
Figure 12. Immunoreactive corticotropes in the pituitary gland in control and genistein treated middle-aged rats. Scale bar 16 μm
Figure 13. The volume density of corticotropes (ACTH cells; A), The volume of ACTH cells (B), plasma concentration of ACTH (C) in control and genistein treated middle-aged male rats. The values are means ± standard deviation, n = 6 animals per group; * p<0.05 vs. control
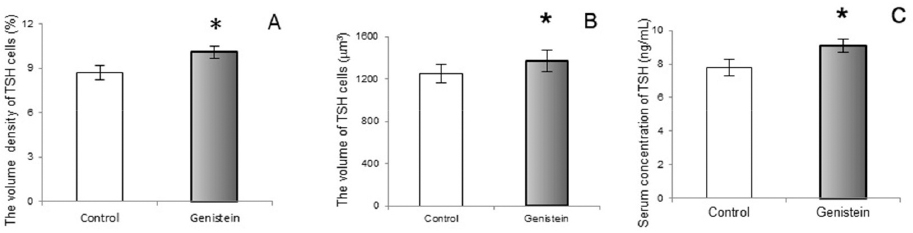
Following genistein treatment pituitary TSH cells were larger and more numerous in middle-aged rats. Also, their relative cellular volumes, as well as the relative volume density per pituitary, unit volume and serum TSH level significantly increased (Fig. 14 and Fig. 15). These changes are probably due to the release from the negative feedback effect of thyroid hormones. (88). Also, Modaresi et al. (89) obtained increased TSH level in mice feeding on soybean.
Figure 14. Immunoreactive thyrotrophs in the pituitary gland in control and genistein treated middle-aged male rats. Scale bar 16 μm
Figure 15. The volume density of thyrotrophs (TSH cells;A), The volume of TSH cells (B), Serum concentration of TSH (C) in control and genistein treated middle-aged male rats. The values are means ± standard deviation, n = 6 animals per group; * p<0.05 vs. control
As with many other compounds, there are many benefits and weaknesses associated with genistein exposure. Unequivocally, genistein contributes to regulation of either morphological and/or functional features of all pituitary hormone producing cells. In which direction HP cells will undergo processes of remodeling following genistein exposure depends on gender, age, hormonal status and dose. Consumers should be aware that genistein is an endocrine disrupting compound and should make dietary choices accordingly.
©2017 Trifunović S. This is an open-access article published under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This research was supported by grants from the Ministry of Education and Science of the Republic of Serbia (No.173009).
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 41, Issue 1, Pages 5-19, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.1515/macvetrev-2017-0027, 2018