Mac Vet Rev 2018; 41 (2): 115 - 122
 10.2478/macvetrev-2018-0013
10.2478/macvetrev-2018-0013
Received: 28 November 2017
Received in revised form: 27 January 2018
Accepted: 12 February 2018
Available Online First: 31 March 2018
Published on: 15 October 2018
Keywords: nucleolus precursor body, oocyte, embryo, transcriptomics, epigenetics
The oocyte acquires competences for sustaining embryonic development during its growth phase (1). We have demonstrated that when bovine and porcine ovarian follicles reach a diameter of about 3 mm, the oocyte has completed its growth. At that time the nucleolus is inactivated to a sphere of densely packed material (nucleolus precursor body; NPB) and oocyte transcription ceases (2, 3).
We have also clearly visualized how the NPB dissolves during the final oocyte maturation and how it again assembles in the two pronuclei of the one-cell fertilized egg, the zygote (4, 5, 6). The NPBs are anlages where functional ribosome synthesizing nucleoli develop a few cell cycles later, when transcription of the ribosomal RNA genes is activated during the major embryonic genome activation (7, 8). Recent studies in pig and mouse, where the oocyte’s NPB was removed (enucleolation) or exchanged by micromanipulation, demonstrated that the NPB is completely essential both for completion of the oocyte’s meiosis (9) and for embryonic development (10).
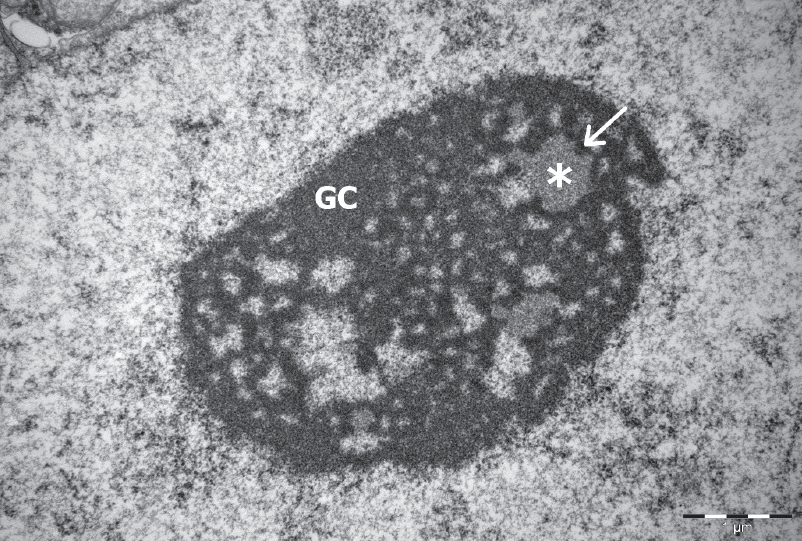
Figure 1. An electronogram of NPB in fully grown porcine oocyte. Scale bar: 2.0 μm
The typical, transcriptionally active nucleolus is composed of fibrillary centers (FC), a dense fibrillar component (DFC) and a granular component (GC) (11). This type of nucleolus is called fibrillo-granullar nucleolus. It has been shown that the sub-compartments of the nucleolus host different proteins involved in transcription, processing and the assembly of pre-ribozomal units. Thus, FC and DFC contain e.g. upstream binding factor (UBF), RNA polymerase I, promotor selectivity factor (SL-1) and topoisomerase I (12, 13). It is well accepted that rDNA transcription occurs at the interface between the FC and the DFC (14). Consequently, the nascent rRNA is located mainly in the FC and the inner portion of the DFC, along with proteins directly involved in the early processing of the rRNA (e.g. fibrillarin). Other proteins (e.g. nucleolin and nucleophosmin) involved in the later steps of rRNA processing and formation of ribosome subunits are located in the DFC, respectively.
It has been fully accepted, for a long period of time, that the main function of the nucleolus in normal cycling cells is the biosynthesis of the ribosomes. However, recent observations in different models have shown that nucleolus is involved also in the processes regulating mitosis, cell cycle progression, proliferation, stress response, pluripotency, cellular reprogramming, etc. Therefore, it is now well accepted that the nucleolus is a multifunctional structure (15). Indeed, in yeasts and somatic cells the nucleolus, or at least material from it, is involved in cell cycle regulation with special emphasis on the transition from metaphase to anaphase (16). In yeast, inactive Cdc14 (protein phosphatase) is localized in the nucleolus during the interphase, but in the anaphase Cdc14 is activated, released from the nucleolus and controls the dissolution of cohesins, a protein complex that regulates the separation of sister chromatids during mitosis and meiosis. In somatic cells, NuSAP (nucleolar-spindle associated protein) is localized in the nucleoli in the interphase, but in the M-phase it is detected in central spindle microtubules. Depletion of NuSAP results in aberrant mitotic spindles and abnormal chromosome segregation and cytokinesis.
It was well documented that upon the activation of oocyte growth, functional nucleolus is established. However, at the end of the growth phase the nucleolus is disassembled, which associates with a gradual extrusion of fibrillar centers to the surface of the nucleolus. During this process of nucleolus re-organization, rRNA synthesis ceases and the nucleolus is, to a high degree, disassembled at the end of the oocyte growth phase (2, 17, 18).
It has been suggested that the nucleoli of fully-grown oocytes are not involved in rRNA synthesis (8). However, in our earlier observations we were able to detect a slow but clear accumulation of [5-3H]uridine-labeled RNA (i.e. non-pre-rRNA) in NPBs of pig antral (19) and mouse preovulatory follicles (20). According to these observations it appears that most of the slowly labeled RNA, which can at least in part represent snRNAs and snoRNAs, occurs in NPB of fully grown oocytes, and that maternal nuclear RNA synthesized by the oocyte concentrates in the NPB at a higher level than elsewhere in the nucleus. However, the accumulation of slowly labeled RNA in preovulatory porcine oocytes, as observed in mouse and human (21), has not been studied so far. Moreover, it has been observed recently, that the remodeling of chromatin into the configuration observed in pre-ovulatory oocytes is not strictly required for global transcriptional repression in mammalian oocytes (22).
In addition, other results obtained in mouse indicate that the NPB might have a structural role in the transcription by other RNA polymerases than RNA polymerase I. It was shown that although the transcriptional activity of both RNA polymerase I and II decreases as the oocytes reach their full size, residual transcriptional activity can still often be found in close vicinity of the NPB in fully-grown mouse oocytes. This activity was further confirmed by RNA fluorescence in situ hybridization with a generic probe composed of non-coding sequences and indirectly, also, by the localization of the RNA-binding protein Sm antigen (unpublished results). The above mentioned residual transcriptional activity was assigned to RNA polymerase I by using transcriptional inhibitor alpha-amanitin (21). However, recent results indicate that as the oocytes undergo transcriptional silencing, RNA polymerase II is redistributed and localizes namely in the vicinity of the NPB (23). Thus, these results point to the possible function of the NPB in transcriptional activity in general.
Figure 2. An electronogram of nucleolar organization in porcine embryo at the morula stage. The three nucleolar components are visible: the fibrillar centres – FC (asterisks), the dense fibrillar component – DFC (arrow), and the granular component (GC). Scale bar: 1.0 μm
The transcriptional activity/silencing was partly studied in mouse. In this species, it was found that transcriptional silencing is brought about by extensive degradation of transcriptional machinery components (23). However, it is not clear whether such mechanism applies to oocytes of all species and thus represents a universal mechanism. Indeed, our preliminary results in porcine oocytes indicate that the mechanism might be more dependent on the phosphorylation status of the components of the transcriptional machinery. Thus, further experiments are critically needed to elucidate the mechanism of transcriptional silencing that occurs during oocyte growth and NPB formation.
In addition to the disputable requirement for the NPB to control meiotic progression and convey essential matters of life to the zygote, it is still an unresolved enigma what type of biomolecules is included in this message. In somatic cells, proteomics has demonstrated a flux of 489 endogenous nucleolar proteins in response to metabolic inhibitors (24). The authors concluded that there is no unique complete proteome for the nucleolus, but rather overlapping sets of proteomes relevant to different cell states or conditions. Further, a few candidate proteins (i.e. NPM2, NCL, SSRP1 and NOLC1) have been found in mouse oocyte NPB (25), together with other key nucleolar proteins (26). Likewise, the somatic nucleolus is rich in small RNAs including microRNAs (miRNAs) (27). Modification of rRNAs is supported by numerous small nucleolar RNAs (snoRNA) with miRNA-properties (28). However, the functional constituents in the oocyte NPB have yet to be identified.
Hours after fertilization, pronuclei appear in the zygote, and NPBs, ultrastructurally resembling the oocyte’s NPB, emerge in the pronuclei (4). At a species-specific time of development, these bodies serve as anlage for the development of active ribosome synthesizing nucleoli. The NPBs, however, have multiple other functions besides being indispensable in organizing and regulating major and minor satellite repeats in the nuclei. Indeed, a recent study by Fulka et al. shows that mouse embryos obtained from enucleolated oocyte (i.e. after removal of the nucleolar sphere from the oocyte) were unable to achieve development beyond the 2-cell stage (10). In these embryos, ribosomal RNA (rRNA) synthesis and pre-rRNA processing were still taking, but not the transcription of the major and minor satellite repeated sequences that is known to be required for proper reorganization of pericentromeric heterochromatin and for early mouse development (29, 30). Moreover, absence of NPBs leads to an abnormal deposition of DAXX (an H3.3 histone chaperone) and consequently, to an abnormal remodeling of the satellite sequences. It is therefore hypothesized that NPBs should contain at least proteins involved in the proper extensive remodeling of pericentric heterochromatin that occurs during the two first cell cycles (31, 32). We assume that NPB could also contain (maternally inherited) RNA and proteins involved in the epigenetic mechanisms which permit proper gene expression in time and space during development.
After establishing the method of enucleolation, the field was opened again for new adventures. The method of enucleolation was described in our last review (39). The studies of Ogushi et al. (10) in pig and mouse, where the oocyte’s NPBs were removed and even exchanged by micromanipulation, demonstrated that the NPB may have effects on oocyte maturation and progression of meiosis and that it certainly is crucial for embryonic development. With respect to meiosis, the first set of studies demonstrated that the NPB was not essential for progression of meiosis in the pig (10). However, later investigations by the same authors disputed this. When growing oocytes are treated by Actinomycin D, the nucleolus compacts into a NPB, and when such NPBs are exchanged with NPBs of fully grown oocytes, the latter lost the ability to progress through meiosis (9). Hence, the NPB may be involved in cell cycle control during the progression of meiosis in porcine oocytes. With respect to embryonic development, the first sets of data were clear: Oocytes that had their NPB removed did not sustain embryonic development. However, if another nucleolar sphere was transplanted to replace the lost one, embryonic developmental capacity was restored. Interestingly, if a nucleolus from a somatic cell or even an embryonic stem cell was used for the transplantation, the oocyte again did not sustain embryonic development. The latter fact indicates that the substances of the oocyte’s NPB which conveys to the conceptus are not merrily nucleolar material, but other biomolecules that are crucial for the embryo to develop. Likewise, enucleated oocytes (cytoplasts) that had their NPBs removed before they underwent final maturation did not support development of cloned embryos after somatic cell nuclear transfer, whereas if nucleolar spheres were introduced into the cytoplast with the somatic cells, developmental capacity was restored. Hence, the oocyte’s NPB is indeed required for conception and further life. Interestingly, later they looked into whether active ribosome synthesizing nucleoli from growing oocytes could substitute the compacted NPB in the fully grown oocyte after micromanipulatory exchange. It turned out that when active oocyte nucleoli are aspirated, chromatin remains attached to them. Transplantation of such active nucleoli were unable to sustain embryonic development, whereas transplantation of NPBs from Actinomycin D-treated oocytes (resulting in nucleolar compaction) were able to restore developmental capacity. Hence, the nucleolus develops the ability to compact into the life-giving NPB already during oocyte growth. Controversially, the same group of authors recently demonstrated that the NPB is dispensable at least in mouse embryonic development (33). If this is the case in other species as well, then this needs to be substantiated.
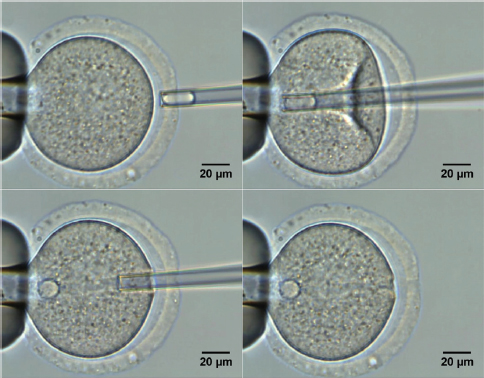
Figure 3. Nucleoli injication into the enucleolated mouse oocyte (fontot na tekstot pod slika broj 3 da e 10)
Interestingly, we have observed that the protein concentration in NPB originated from mouse oocytes is higher than in pig oocytes (34), while mouse NPBs were sufficient in securing embryonic development in the pig (35). Mouse embryos resulting from enucleolated oocytes (i.e. lacking the NPB) were unable to develop beyond the 2-cell stage (10). In such embryos, ribosomal RNA (rRNA) synthesis and pre-rRNA processing occurred, but transcription of sets of satellite repeats, required for proper reorganization of pericentromeric heterochromatin (29, 30), was lacking. Hence, we believe that the NPB contains substances involved in the epigenetic remodeling of chromatin that occurs during the first embryonic cell cycles (25, 31, 32) and which sets the stage for proper gene expression in time and space during embryonic development. Hence, the oocyte’s NPB is indeed required for conception and further life.
Mammalian oogenesis, oocyte maturation and early embryo development are distinct processes, each carefully coordinated, and bound by a common thread: the need to supply an appropriately programmed genome that permits proper gene expression in time and space. In addition to transcription factors binding to promoters, regulation of transcription early in development is achieved via various epigenetic mechanisms. The epigenetic regulation occurs on the transcriptional (DNA methylation, histone modification, nucleosome positioning), as well as on the post-transcriptional level (miRNA). The interplay between these processes is crucial for maturation/differentiation in growing oocytes, global genome silencing during the peri-fertilization period, and the determination of proper timing and the extent of de-differentiation and embryonic genomic activation in pre-implantation embryos.
As shown previously (8), porcine pre-implantation embryos do not display transcriptional activity, indicating the presence of extensive silencing marks on chromatin. The reprogramming of epigenetic memory, basic metabolic processes as well as embryonic genome activation are thus fully dependent on maternally inherited material, mRNA and rRNA, gradually being used by the embryo. However, up to now, nothing is known about the basic essential factors responsible for proper use of these RNAs. One of the most studied epigenetic modifications is DNA methylation. The level of DNA methylation of development-related genes (e.g. Oct4, Nanog, Sox2, e-cadherin, etc.) decides the silencing and/or expression of these genes and undergoes dramatic changes between oocyte maturation (hypermethylation) and early embryogenesis (hypomethylation). Moreover, the proper maintenance of parentally determined DNA methylation imprints during germ cell maturation and the postfertilization period is crucial for a proper development of concepti. Beside DNA methylation, recent data on miRNA epigenetic regulation in mouse (36) suggests that endogenous small interfering RNA (endo-siRNA) will be the key players in determining the early pre-implantation development in the porcine zygotes and embryos (37). Together with the miRNA in the early peri- and postfertilization period, specific histone modifications are also responsible for successful genome activation. Histone modifications are under the control of specific histone acetyl transferases (HAT) and histone deacetylases (HDAC). The activity of these regulates the overall acquirement of gene silencing (H3K9me3, H3K27me3) and/or active marks (acH4K16, H3K4me3). Therefore, the possible aberrations in the presence and activity of these enzymes may have detrimental effect on early embryogenesis and lead to developmental arrest of embryos (38). However, if and to which extent the epigenetic changes influence an early embryonic development in embryos produced by nucleolotransfer has not been evaluated so far.
It has been shown than NPB is of utmost importance for the developmental competence of the fertilized egg and developing embryo. Hence, the nucleolar sphere contains matters that convey developmental competence to the conceptus. It is, however, unknown what this enigmatic structure contains and in what way it contributes to embryonic development. Therefore, the content of the NPB has to be analyzed by advanced molecular approaches, and the developmental competence of the NPB has to be tested by advanced micromanipulation where NPBs are transplanted from one egg to another. This will bring new knowledge about potential functions of NPB in early development of the embryos used by assisted reproductive technologies and biomedicine.
©2018 Benc M. This is an open-access article published under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This work was supported by Slovak Research and Development Agency under the contract No. APVV-14-0001, and also by the projects CZ.02.1.01/0.0/0.0/15_003/0000460, VEGA 1/0022/15, VEGA 1/0327/16 and Research Center AgroBioTech built in accordance with the project Building Research Centre „AgroBioTech” ITMS 26220220180. JF Jr. is supported from GACR 17-08605S.
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 41, Issue 2, Pages 115-122, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.2478/macvetrev-2018-0013, 2018