Mac Vet Rev 2019; 42 (1): 5 - 13
 10.2478/macvetrev-2018-0023
10.2478/macvetrev-2018-0023
Received: 09 January 2017
Received in revised form: 15 April 2018
Accepted: 07 May 2018
Available Online First: 03 November 2018
Published on: 15 March 2019
Keywords: andropause; testosterone; gonadotropic cells; immunohistomorphometry; optical density
The complex phenomenon of ageing in males, which implies a temporal, psychophysiological and social dimension, is progressive and gradual in its essence, and culminates in a point of entry into andropause (1, 2, 3) i.e. the specific multi-symptomatic syndrome that is substantially characterized by deregulation of the hypothalamic-pituitary-gonadal (HPG) axis (4). Free testosterone deficiency, which grounds the hormonal milieu of the ageing male, represents the consequence of numerous and multilevel alterations. When it comes to testicular Leydig cells, lowered activity of a certain steroidogenic enzymes or increase in cyclooxygenase-2 activity and its tonic inhibition of steroidogenic acute regulatory gene expression are observable (5). Also, some studies with aged rats suggested the existence of hypothalamic-pituitary dysfunction characterized by a reduction in the gonadotrophin releasing hormone (GnRH) impulse from the hypothalamus and a decrease in the amplitude of the luteinizing hormone (LH) secretory pulse (6, 7). In conjunction with these results, Elmilinger et al. (8) reported a significant increase for pituitary follicle-stimulating hormone (FSH) and luteinizing hormone (LH) serum levels with ageing in men, which may indicate the long-term feedback effect of low testosterone (9).
Testosterone supplementation in andropause seems to be an approach of choice in the symptomatic approach (10, 11), although a considerable monitoring for efficacy and safety is advised (12). Furthermore, circulating levels of LH and FSH were shown to be significantly decreased during dihydrotestosterone treatment of ageing men (13), while applied testosterone supressed the pulsatile LH and FSH secretion in castrated rats (14). Testosteron regulates the gonadotrope cell function by a negative feedback effect, achieved at the level of the hypothalamus and pituitary gonadotropic cells directly, where the expression of androgen receptors was found (15). We hypothesised that the immunohistomorphometric properties of pituitary gonadotropic cells (FSH and LH producing, the specific operative units of HPG axis) may change after testosterone application to ageing male rats, so the aim was to visualize a phenomenology of the potential change by using available microscopic methodology.
Therefore, the immunohistomorphometric properties of the FSH and LH cells were in our focus, in an animal model of the andropause. Orchidectomized middle-aged rats, in which both testicular testosterone and estradiol are eliminated from the hormone milieu, were used in order to mimic the andropause. In parallel, this experimental set up fits into an established approach for examining the potential effects of synthetic steroids/steroid-like compounds on endocrine homeostasis (16 - 19).
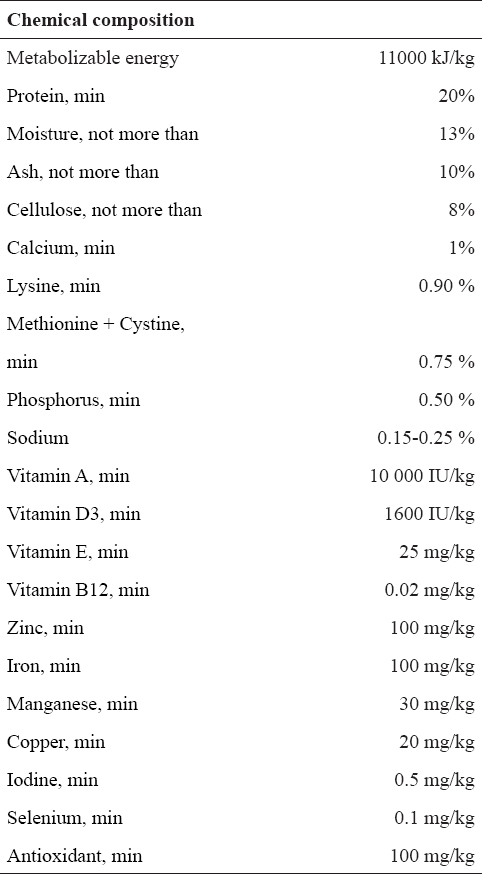
The experiments involved 16-month-old (middle-aged) Wistar rats. They were bred in the Institute for Biological Research “Siniša Stanković”, Belgrade, Serbia; housed one per cage and maintained under constant laboratory conditions (22±2ºC, 12-12 h light-dark cycle), with free access to food (standard diet; the chemical composition given in Table 1) and water.
Table 1. Chemical composition of the rat diet used in the experiment
At the age of 15 months, experimental animals were randomly bilaterally orhidectomized (ORX; n=16 animals) under Ketamine anesthesia (15mg/kg b.m.; Richter Pharma, Wels, Austria). The recovery period lasted two weeks. Orchidectomized rats were then divided into two groups of eight animals (n=8) each. The first group was subcutaneously treated with testosterone propionate (Fluka Chemie AG, Buchs, Switzerland; ORX+T), in a dose of 5 mg/kg b.m. (19, 25-26) every day except on Sundays, for 3 weeks. The final injected volume was 0.5 ml per animal. The second orchidectomized group (ORX) was given the same volume (0.5 ml) of vehicle (sterile olive oil) alone, and they served as controls. Body mass, from these two groups after the treatment, was measured. All the animals (n=16) were sacrificed by decapitation under ether anesthesia (ether ad narcosis Ph. Iug. III., Lek, Ljubljana, Slovenia), 24 h after the last injection.
All experimental procedures were in compliance with Directive 2010/63/EU on the protection of animals used for experimental and other scientific purposes and was approved by the Ethical Committee for the Use of Laboratory Animals of the Institute for Biological Research “Siniša Stanković”, University of Belgrade, Serbia.
Blood was collected from the trunk and separated sera samples of all animals were stored at the same time at -70°C until assayed. Serum testosterone concentrations were determined without dilution, using the competitive immunoenzymatic colorimetric method (EIAgen Testosterone Kit; Adaltis Italia S.p.A., Bologna, Italy), which sensitivity is 0.01 ng/ml, with an intra-assay coeficient of variation (CV) of 6.2%. Serum estradiol concentrations were measured without dilution, using the electrochemiluminescence immunoassay (ECLIA, Roche, Basel, Switzerland), which low detection limit is 5.0 pg/ml, with an intra-assay CV of 6.2%.
After sacrificing, excised and weighed pituitaries were fixed in 4% paraformaldehyde for 24 h, dehydrated in a series of increasing concentrations of ethanol, enlightened in xylol and embedded in paraplast. Serial, 5-µm thick pituitary tissue sections were obtained using LEICA RM 2125RT rotational microtome (Leica Biosystems Nussloch GmbH, Nussloch, Germany), then deparaffinised in xylol and rehydrated in a series of decreasing concentrations of ethanol. For the light microscopy analysis, gonadotropic- (FSH- and LH- producing) cells were immunolabeled using the peroxidase-antiperoxidase complex (PAP) method (20, 21). After rehydration of the pituitary sections, the endogenous peroxidase activity was blocked by the incubation in 9 mmol (0.3%) hydrogen peroxide solution in methanol, for 15 min at 24°C. Before the application of the specific primary antisera, nonspecific background staining was prevented by incubation of the sections with nonimmune, i.e. normal swine serum, diluted (1:10) with phosphate buffered saline (PBS) pH 7.4, for 45 min at 24°C. Pituitary sections were then overlaid with the appropriate dilutions of the specific primary antibodies, namely polyclonal anti-rat βFSH (1:300) and βLH (1:500), for 24 h at 4°C. Antisera to rat βFSH (NIDDK-anti-rBeta FSH-IC1, Lot#AFP-7798 1289P) and βLH (NIDDK-anti-rBeta LH-IC, Lot#AFP-C697071P) were obtained from Dr. A. F. Parlow, National Hormone Peptide Program (NHPP), Harbor-UCLA Medical Centre, Carson, CA, USA. The specificities of these antisera were assessed by the NIDDK. After washing in PBS, the sections were incubated with secondary antibody polyclonal swine-anti-rabbit IgG/HRP (Dako A/S, Glostrup, Denmark). Binding sites were visualized using 0.05% diaminobenzidine (DAB, Serva, Heidelberg, Germany) and 0.03% hydrogen peroxide in 0.2M TRIS-HCl buffer, pH 7.4. The sections were counterstained with hematoxylin and mounted in Canada balsam (Molar Chemicals KFT, Budapest, Hungary). For the negative control pituitary sections, the primary antibody was omitted and replaced by PBS.
Digital images of the pituitary gland immunolabeled sections were taken using a LEITZ DM RB light microscope (Leica Mikroskopie & Systems GmbH, Wetzlar, Germany), a LEICA DFC320 CCD camera (Leica Microsystems Ltd., Heerbrugg, Switzerland) and the Leica DFC Twain Software (Leica, Germany).
Morphometrical measurements were performed as described previously (16-18). Namely, two pituitary sections (5-µm thick) from the superior, three from the middle and two from the inferior part (seven horizontal sections) of the rat pituitary glands were analysed. The point counting method was used at an overall magnification of x1000. The M42 multipurpose test grid, inserted into the ocular of a Zeiss light microscope (Jena, Germany), was randomly positioned on the pituitary section at the beginning of counting. Counting was carried out on the following 50 test fields per section. Average values were calculated per pituitary i.e. per animal (7 sections, 350 test fields) and eight pituitaries were analysed per group. Cell volume (Vc, μm3) and relative volume density (percentage of immunopositive cells in mm3; VVC, %) were determined for FSH- and LH-immunopositive cells.
The following parameters were counted:
Pn - number of points hitting on nuclei of immuno-histochemically labeled cells inside the test field
Ptc - number of points hitting on cytoplasm of immuno-histochemically labeled cells inside the test field
Nn - number of immuno-histochemically labeled cell nuclei inside the test field.
The formula for calculating the cell volume was:
Vc = 1/NV , where
NV is the numerical density of FSH and LH cells. Nuclei volume density (VVn) provides information about nuclei attendance in the estimated cells and is calculated as follows:
VVn = ΣPn/ΣPtc
Since rat FSH and LH cells are mononuclear, NV corresponds to the number of cells per cubic millimetre, according to the formula:
NV = (k/β) (NA3/2 / VVn½)
On the basis of earlier karyometric studies (22), the shape coefficient β for pituitary cells was estimated to be 1.32, where k is a factor related to cell distribution according to their size (in the case of FSH and LH cells its value is 1) and NA is the number of cells in the plane of the pituitary tissue section. NA is calculated as follows:
NA = ΣNn/ΣPtc·a ,
where a represents the rhombic area belonging to every point of the test system and is calculated using the formula:
a = d2 31/2/2 ,
where d is the test line length in the test system employed.
Relative volume density (VVC) is calculated as the ratio of the sum of Pn and Ptc (Pn+Ptc) and the total number of points in the test system. Since a test system with 42 points was used and parameters were calculated using 50 test fields, the definite formula was:
VVC = (Pn+Ptc)/ 50·42
The Windows based ImageJ program (Image J, version 1.50f) was used for the analysis of optical density that reflects the gonadotropic cell hormonal content. Namely, 30 unbiasedly captured images (the microscopic tool has already been described: 2088 x 1550 pixels, 63x objective magnification) per gonadotropic cell (FSH or LH) specific-immunostained pituitary per animal were analysed. Initially, the spectral deconvolution method of 3,3′-diaminobenzidine tetrahydrochloride (DAB)/Hematoxylin color spectra was performed, using optimized optical density vectors of the color deconvolution plug-in for adequate separation of the DAB color spectra. To determine the optical density for the red, green and blue (RGB) channel of Hematoxylin and DAB, we kept the protocol previously described by Ruifrok and Johnston (23) and Varghese et al. (24). Since the optical density is proportional to the concentration of the stain in gonadotropic cells, the amount of stain present is a factor determining the optical density (OD) at a wave length specific to the stain, according to the formula:
OD = – log10 (IC/I0.C),
IC is the intensity of detected light after passing through a specimen and I0.C refers to the intensity of light entering the specimen.
STATISTICA® version 7.0 (StatSoft, Inc) was used for the statistical analysis. Hormonal and morphometric data obtained from each group were statistically evaluated by the Student´s t-test. A probability value of 5% or less was considered statistically significant. The data are presented as means ± SD.
Data for the body mass and absolute/relative pituitary weights are summarized in Table 2. It is obvious that orchidectomy, as well as subsequent treatment with testosterone did not cause statistically significant changes in the measured parameters.
Table 2. Body mass, absolute and relative pituitary weights in orchidectomized (ORX) and testosterone treated orchidectomized (ORX+T) middle-aged rats
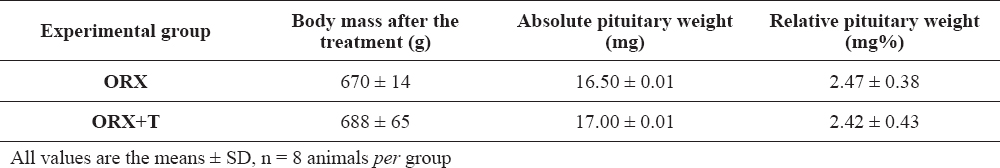
The serum hormonal levels are shown in Figure 1. In ORX+T group, circulating testosterone and estradiol were significantly (p<0.05) increased by 24 and 2.7 fold respectively, compared to the ORX group values (Figure 1A and B).
Figure 1. Serum level of testosterone (A) and estradiol (B) in orchidectomized (ORX) and testosterone treated orchidectomized (ORX+T) middle-aged rats. All values are provided as the mean ± SD, n = 8 animals per group; *p<0.05 vs. ORX rats
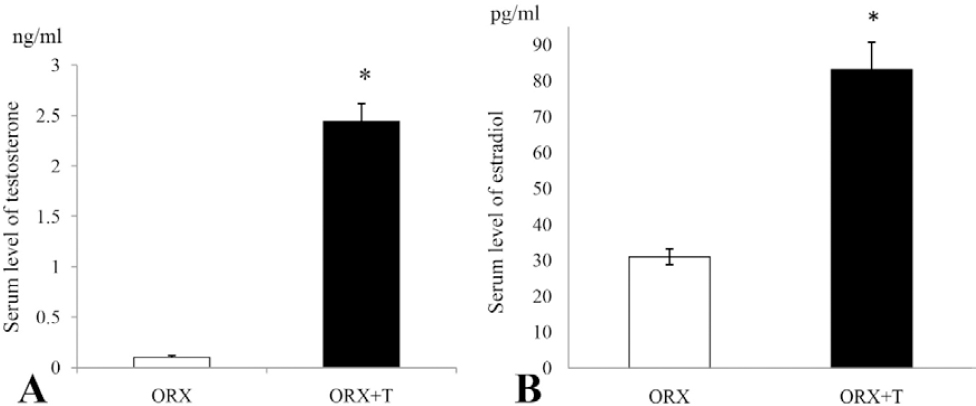
Gonadotropic cells (FSH and LH) in ORX rats were widespread throughout the pars distalis, in groups or as single cells often in close contact with blood capillaries. They were hypertrophic, predominately oval in shape, with prominent eccentrically located nuclei. Strong immunopositivity of FSH and LH cells was observed. After the treatment with testosterone, gonadotropic cells were significantly smaller in size and less numerous (Figure 2A and B).
Figure 2. SH- (A) and LH- (B) immunoreactive cells in the pituitary pars distalis from orchidectomized (ORX) and testosterone treated orchidectomized (ORX+T) middle-aged rats (objective magnification 40x, bar = 25μm)
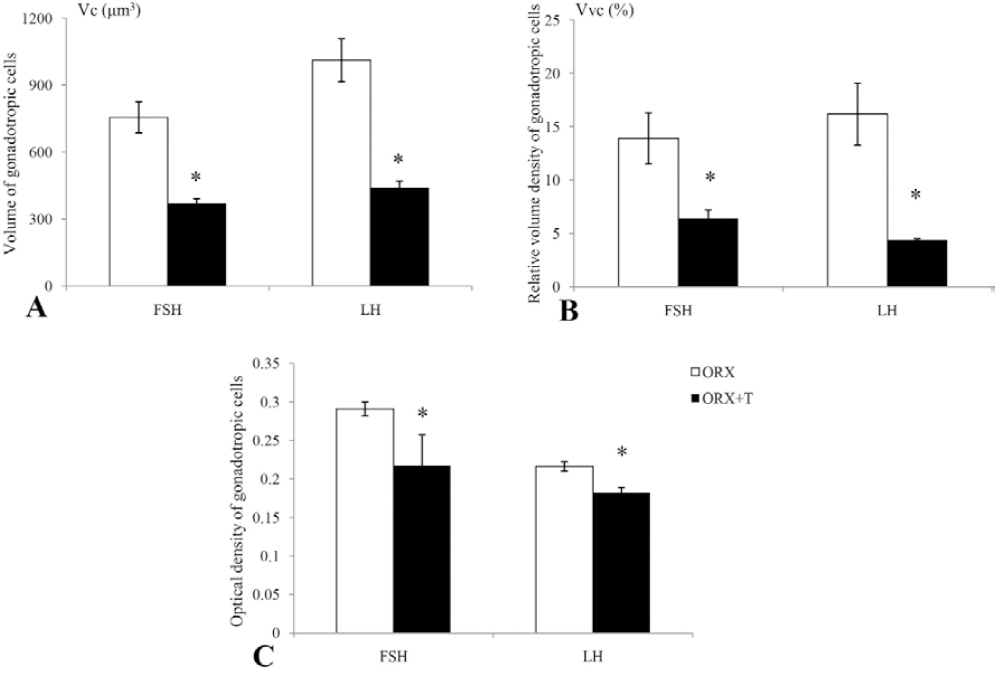
The morphometric and optical density parameters observed for the FSH and LH cells in this study are represented in Figure 3. Namely, in ORX+T rats the volume of FSH and LH cells (Vc) was significantly (p<0.05) decreased by 51.3% and 56.6% respectively, in comparison with the ORX animals (Figure 3A). The relative volume density (Vvc) of FSH and LH cells in ORX+T animals was significantly (p<0.05) decreased by 54.0% and 72.8% respectively, compared to the ORX group (Figure 3B). Optical density of FSH and LH cells after orchidectomy and testosterone treatment was significantly (p<0.05) decreased by 25.7% and 16.2% respectively, in comparison with ORX rats (Figure 3C).
Figure 3. (A) Cellular volume (Vc, μm3), (B) relative volume density (Vvc, %) and (C) optical density of FSH and LH cells in orchidectomized (ORX) and testosterone treated orchidectomized (ORX+T) middle-aged rats. All values are provided as the mean ± SD, n = 8 animals per group. *p<0.05 vs. ORX rats
In the present study, we investigated the effects of testosterone application on the immunohistomorphometric properties of pituitary FSH and LH cells in a rat model of the andropause, adequately represented with orchidectomized middle-aged rats. By performing orchidectomy, any testicular inputs on the HPG axis were avoided, which defines the optimal space for examining the potential effects of synthetic steroids on the endocrine homeostasis (19, 25, 26). The dose of testosterone employed (5 mg/kg b.m.) has already performed some bone protection and stress hormone secretion-related beneficial effects in the same male rats (19, 25, 26). Our aim was to visualize a phenomenology of changes at the level of specific HPG axis operative units, upon application of a beneficial dose of testosterone to andropausal rats, by exploiting the available microscopic methodology.
In our experiment, applied testosterone expectedly increased the serum concentration of testosterone, but raised the circulating estradiol as well. It seems that the considerable aromatase activity (converts androgens into estrogens) is displaced outside the testes in andropausal rats, given that Leydig cells as the important source of aromatase (27) are surgically eliminated herein.
In orchidectomized rats, gondotropic (FSH and LH) cells are hypertrophied and enlarged, which could be explained by increased stimulation by GnRH (28). Increased serum levels of testosterone and estradiol in our study led to the recovery of hyperstimulated gonadotropes, confirmed at the histological level by detection of smaller and less numerous FSH and LH cells. Also, the volume, volume density and optical density (reflecting the hormonal content) of gonadotropic cells were significantly reduced. Our results confirmed that sex steroids regulate gonadotropic cell function by negative feedback effect. At the level of hypothalamus, sex steroids provide a recovery of the GnRH content in ORX male mice and indicate that androgen and estrogen signaling pathways can regulate the GnRH content in castrated males (29). Expression of androgen and estrogen receptors in FSH and LH cells in rats suggests a direct action of sex steroids at the gonadotropic cell level (15, 30). Whether they act at the hypothalamic or pituitary levels or both, testosterone and estradiol suppress FSH and LH cell function (29).
In conclusion, we have demonstrated that the exposure of orchidectomized rats to testosterone causes the reduction of volume, relative volume density and optical density of gonadotropic cells, while it increases serum concentrations of testosterone and estradiol. These changes indicate a potent effect of synthetic testosterone on the function of HPG system and endocrine homeostasis in males.
© 2018 Ristić N. This is an open-access article published under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
This work was supported by the Ministry of Science, Education and Technological Development of the Republic of Serbia, Grant number 173009.
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Macedonian Veterinary Review. Volume 42, Issue 1, Pages 5-13, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.2478/macvetrev-2018-0023, 2018