Abstract
It is well known in cattle that reproductive disorders are intimately associated with low or high body condition score (BCS). However, little is known concerning the relationship between BCS and oxidative stress, particularly in the reproductive tract. Thus, the aim of the present study was to investigate the oxidative stress biomarkers according to cows BCS values in the plasma, ovaries, oviductal, follicular and uterine fluids. The study was conducted on 58 cows classified in four classes (1, 2, 3 and 4) of BCS varying from1-1.5, 2-2.5, 3-3.5 and 4-4.5, respectively. The genital tracts and plasma were collected from slaughtered cows and processed within 5h after slaughtering. The number of ovarian follicles was measured using ultrasonography and the oxidative stress was assessed by considering total antioxidant status (TAS), catalase activity (CAT), and thiobarbituric acid-reactive substances (TBARS). The results showed that follicles number was significantly affected by BCS; cows with medium BCS (3) presented higher number of follicles than cows with low or high BCS (<3 or >3). Overall, BCS affects significantly the oxidative stress status at different levels of the reproductive tract. Cows with medium BCS (3) showed the best oxidative status than those with low or high values (BCS <3 or >3). In conclusion, the current results suggest that reproductive disorders observed in cows with low or high BCS values could be mediated through oxidative stress affecting consequently the reproductive tract and finally compromising fertility outputs.
Keywords: BCS, oxidative status, follicles number, reproductive tract, reproduction
INTRODUCTION
Body condition score (BCS), reflecting the nutritional status in beef and dairy cows, has been extensively considered as a valuable practical tool in herd management. BCS is significantly related to metabolic status, productive and reproductive performances (
1). BCS measures, by visual inspection and/or palpation, the amount of metabolic energy stored as subcutaneous fat (
2). In dairy cows particularly, BCS has been widely used as a practical method in nutrition management (
3) and remains significantly associated with incidence of metabolic disorders (
4). The importance of BCS in fertility has also been reported (
4).
Insufficient intake of energy has been associated with poor reproduction performance in cows with various health problems (
5). In this respect, Stagg et al. (
6) reported that lower luteinizing hormone (LH) pulse frequency induces prolonged anestrus period in cows that calved with low BCS (<2.5). Similarly, it is shown that cows could be pregnant only with BCS of 2.75-3 and not losing more than 0.5 of BCS unit between calving and first service (
7).
On the other hand, excessive body energy reserves can cause several health problems, (reproductive failure, obese and older heifers at puberty that present difficulties during pregnancy) as well as disorders in the development of the mammary gland. As well, excessive fat reserves in dairy cows have been associated with a greater risk of a lowered milk yield and metabolic disorders (
4).
There is considerable evidence to support that oxidative stress contributes to a number of health disorders. Excessive reactive oxygen spices (ROS) might damage macromolecules including lipids, proteins, polysaccharides and DNA, leading to cell injury, and increased susceptibility to a variety of health disorders including immune dysfunction, sepsis, mastitis, acidosis, ketosis, enteritis, pneumonia, and other diseases (
8). Nevertheless, in cattle, there is real scarce information regarding the association between the oxidative status and BCS. Therefore, the aim of the present study was to investigate the impact of BCS on oxidative status at different levels of the reproductive tract.
MATERIAL AND METHODS
Chemicals
The following reagents: sodium chlorid, trichloroacetic acid, potassium dihydrogen phosphate, dipotassium hydrogen phosphate, DPPH (1,1-diphenyl-2-picrylhydrazyl), ABTS (2,2′-azinobis-(3-ethylbenzothiazoline-6-sulphonic acid diamonium salt), potassium persulfate, hydrogen peroxide, TBA (thiobarbituric acid), sodium hydroxide (NaOH), ethylenediaminetetraacetic acid (EDTA) were obtained from commercial suppliers and used without further purification.
Animals
A total of 58 cows were selected for this study and classified according to their body condition score (BCS). BCS were assigned by experienced veterinarian using a five-point scale method (scale of 1 to 5) as described previously (
9). On the basis of the individual values, four BCS classes were considered: 1 (1-1.5), 2 (2-2.5), 3 (3-3.5) and 4 (4-4.5).
Preparation of plasma and isolation of bovine erythrocytes
Blood samples were collected into heparinized tubes from cows before slaughtering. Blood was centrifuged at 5000g for 10 minutes to separate plasma. Plasma was stored at -20 °C for late use.
Collection of follicular fluid, oviductal fluid, ovaries and uterine fluid
The genital tracts of the studied cows were obtained from slaughterhouse. The tracts were transported to the laboratory in a cooler (4 °C). All samples were collected and processed within 1 hour after slaughtering.
Morphology of the corpus luteum was assessed for each individual cow to estimate the stage of the estrous cycle and the follicles number was determined using an ultrasonography.
Follicular fluid
Follicular fluid was aspirated from each follicle with an insulin syringe, due to the low volume of removed follicular fluid from the different follicles of the two ovaries. Samples were kept on ice until centrifugation for 5 minutes at 5400g to remove blood cells. The clear supernatants were aliquoted and stored at -20 °C until use.
Oviductal fluid
Oviducts were separated from the ovaries and the enveloping connective tissues were removed. Oviductal epithelial cells were recovered in plastic tubes by flushing method; each oviduct was flushed with 1ml of physiological solution (NaCl 0.9%). The cells were lysed in three volumes of distilled water, and then centrifuged at 5000g for 10 minutes; the supernatants were collected and stored at -20 °C until use.
Uterine fluid
The fluid was collected by flushing the uterine cavity with 10 ml of NaCl 0.9%. The fluid was aliquoted and stored at -20 °C.
Ovaries homogenate
The ovaries were homogenized using a blender. The homogenate was lysed in distilled water, and lysate was centrifuged at 5000g for 15 minutes to obtain the supernatant which was used as a whole cell extract.
Measurement of antioxidant activity
Three biochemical markers of oxidative stress were measured including total antioxidant status (TAS), catalase activity (CAT), and the level of lipid peroxidation (TBARS). These biomarkers were measured in the ovaries, oviductal and uterine fluids, follicular fluid and plasma.
Catalase activity
Catalase activity was determined using the method of Aebi (
10) by measuring the degradation rate of H2O2. Briefly, 20 µl of tissue sample were incubated with 1250 µl of PBS then 730µl of H2O2 (50 mmol/l) were added. The decrease in absorbance at 240 nm was measured. Catalase activity was expressed in terms of 1moles of H2O2 consumed/min/ml.
Total antioxidant status
The antioxidant status of tissue samples was evaluated according to the method of Re et al. (
11). This assay is based on the inhibition of the absorbance of the free radical cation from ABTS (2,2’-azinobis (3-ethylbenzthiazoline-6-sulfonic acid)) by antioxidants. Briefly, ABTS was dissolved in deionized water to make a 7 mM concentration solution. ABTS was incubated with 2.45 mM potassium persulfate in order to produce the free radical cation (ABTS°+) and the mixture was allowed to stand in the dark at room temperature for 12–16 h before use. For our study, the ABTS°+ solution was diluted to an absorbance of 0.7 (±0.02) at 734 nm. A volume of 10 µl of each tissue sample was added of 2 ml of diluted ABTS°+ solution. The absorbance reading was taken exactly 6 min after initial mixing. All experiments were carried out in triplicate. Radical scavenging activity was expressed as the percentage of inhibition.
Measurement of lipid peroxidation
Thiobarbituric acid reactive substances (TBARS) were determined following the method of Stocks and Dormandy (
12). Tissue samples were deproteinised by adding trichloroacetic acid (0.5 ml of 30% TCA) and subsequently incubated at 0 °C for 120 min. The mixture was centrifuged at 2500g for 10 minutes at 4 °C and a portion of the supernatant (1 ml) was mixed with thiobarbituric acid (0.25 ml of 1% TBA dissolved in 0.05 mol/l of NaOH) and EDTA (0.075 ml of 0.1 mol/l EDTA) and heated at 95 °C for 15 min then immediately cooled under tap water. The absorbance of the supernatant was determined at 535 nm against a blank that contained all the reagents minus the biological sample. The concentration of TBARS was expressed in absorbance units.
Statistical analysis
All experiments were carried out in triplicate and the data are given as means ± S.E. Calculation of means, S.E, and statistical analysis were performed using Statview 4.02 software (Abacus Concepts Inc., Berkeley, CA, USA). Values of each parameter were expressed as the mean ± S.E. Variables used for comparison purposes were the BCS classes. Differences between treatments were assessed using a one-way ANOVA, followed by posthoc Fisher’s test. Values were considered significant when P < 0.05.
RESULTS
Follicles number
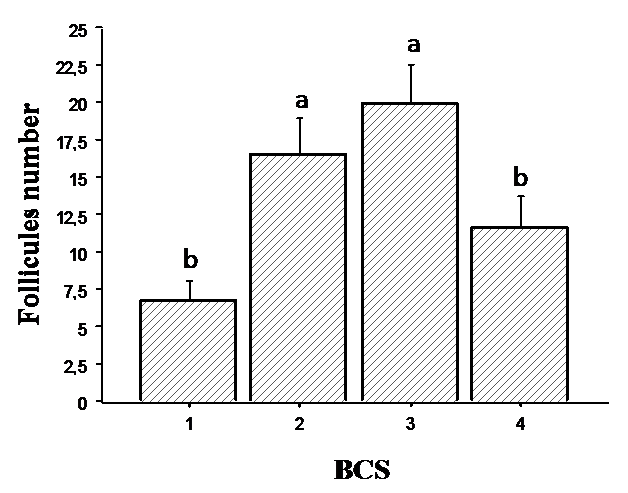
Follicles number detected by ultrasonography according to BCS values is given in
Fig. 1. Cows having BCS 3 and 2 presented the maximum number of follicles with values of 19.87±2.76 and 16.50±2.56, respectively. In contrast, cows having BCS 1 and 4 showed the lowest follicles number with values of 6.75±1.39 and 11.66±2.10, respectively.
Figure 1. Follicles number measured ultrasonographically in relation to body condition score (BCS). Values are represented as mean ± S.E. Different letters indicate significant differences (P < 0.05)
Total antioxidant status
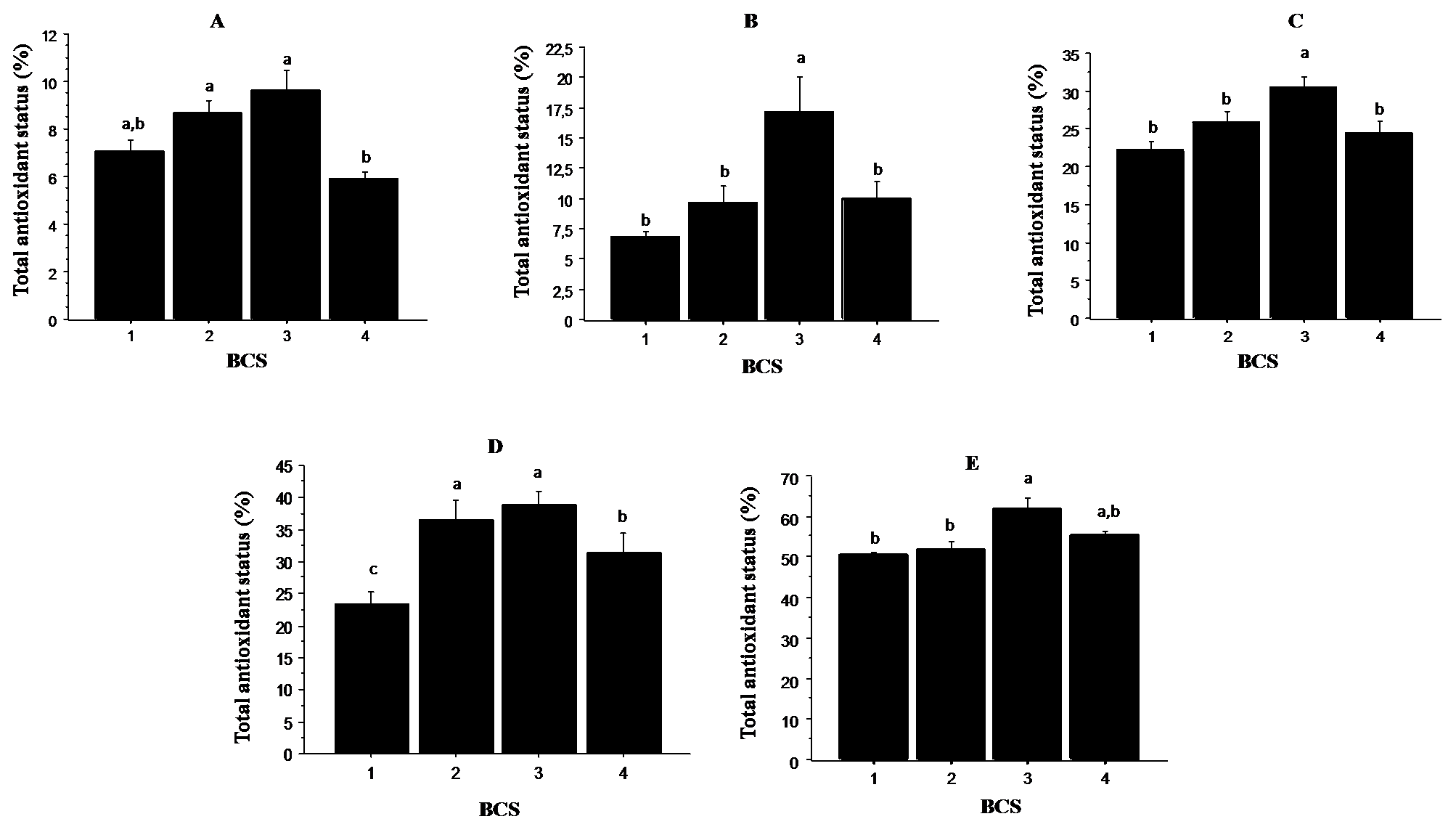
Fig. 2. illustrates the results of the antiradical activity expressed as a percentage inhibition of ABTS. It was found that the total antioxidant status of the oviductal fluid (panel A), uterine fluid (panel B), follicular fluid (panel C), ovaries (panel D) and plasma (panel E) were significantly higher (P < 0.05) in cows with BCS 3. Cows with BCS 1 and 4 presented the lowest total antioxidant status in all the analyzed tissues. The intermediate values were observed in cows with BCS 2.
Figure 2. Relationship between total antioxidant status and BCS in oviductal fluid (panel A), uterine fluid (panel B), follicular fluid (panel C), ovaries (panel D) and plasma (panel E). Values are represented as mean ± S.E. Different letters indicate significant differences (P < 0.05)
Catalase activity
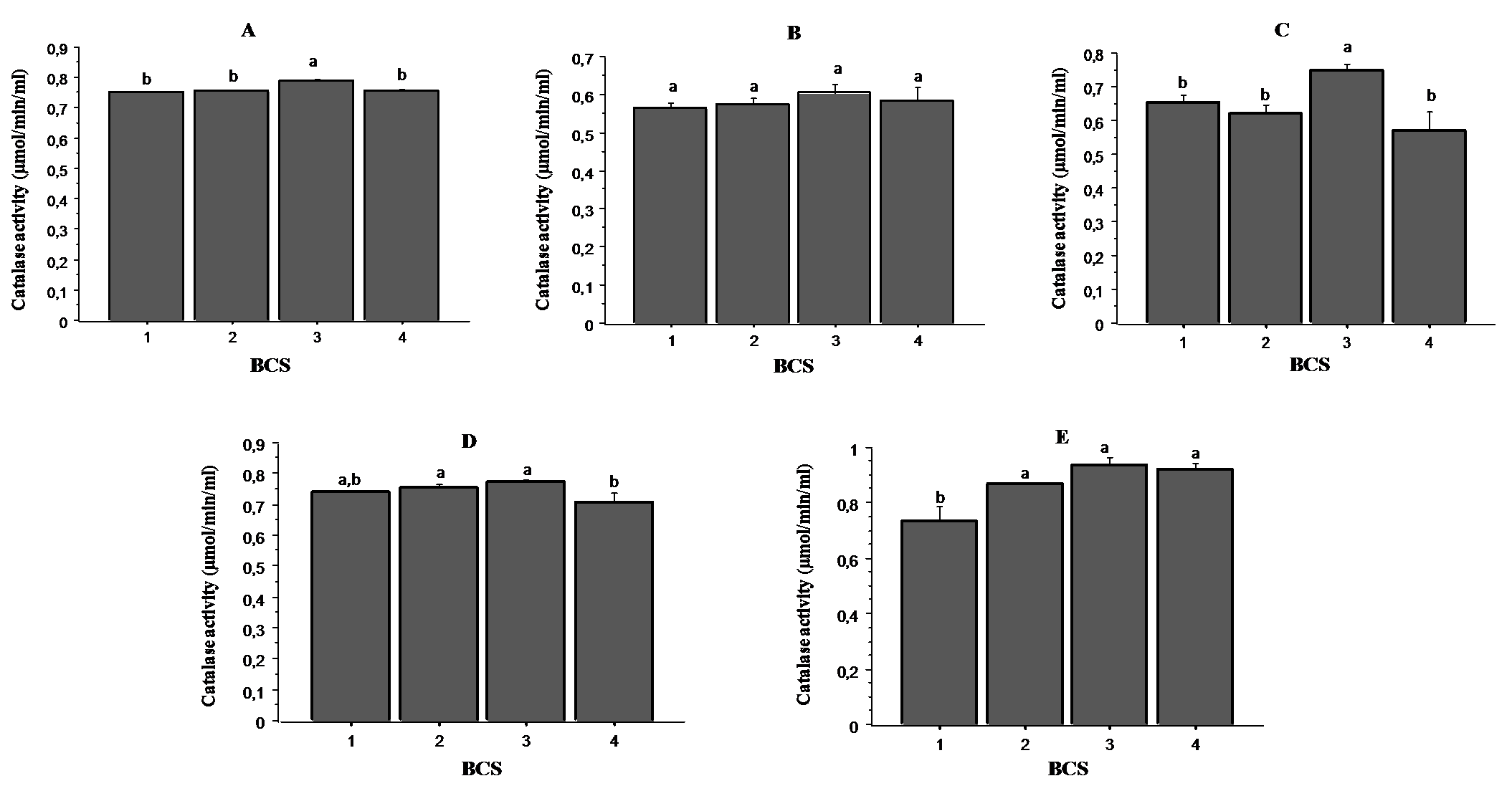
Catalase is an important enzyme in the antioxidant defense system protecting animals from oxidative stress. It catalyzes the conversion of the hydrogen peroxide (H2O2) into oxygen and water.
Fig. 3. shows the results of catalase activity in oviductal fluid (panel A), uterine fluid (panel B), follicular fluid (panel C), ovaries (panel D) and plasma (panel E) according to BCS. The highest activity was recorded in samples from cows with BCS 3, while cows having BCS 4 presented the lowest catalase activity, particularly in the follicular fluids and ovaries. In the plasma samples, cows with BCS 1 showed the lowest activity.
Figure 3. Relationship between catalase activity and BCS in oviductal fluid (panel A), uterine fluid (panel B), follicular fluid (panel C), ovaries (panel D) and plasma (panel E). Values are represented as mean ± S.E. Different letters indicate significant differences (P < 0.05)
Lipid peroxidation
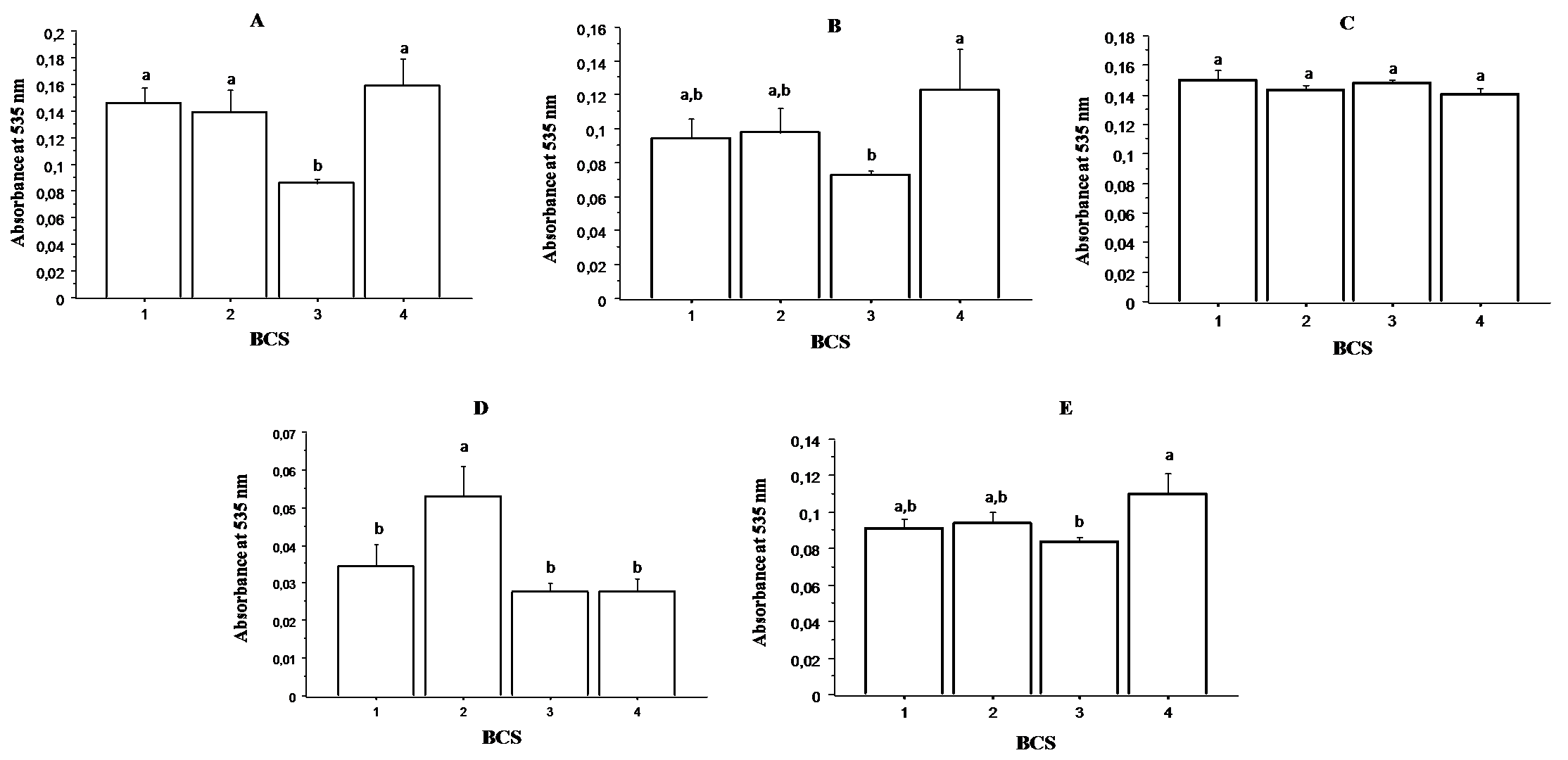
The data are shown in
Fig. 4. No significant differences in lipid peroxidation were observed in the Follicular fluid (panel C) among the cows having different BCS. However, concerning the other analyzed samples (oviductal fluid (panel A), uterine fluid (panel B), ovaries (panel D) and plasma (panel E)), the lowest lipid peroxidation was observed in cows having BCS 3.
Figure 4. Relationship between TBARS and BCS in oviductal fluid (panel A), uterine fluid (panel B), follicular fluid (panel C), ovaries (panel D) and plasma (panel E). Values are represented as mean ± S.E.
Different letters indicate significant differences (P < 0.05)
DISCUSSION
The nutritional status, evaluated trough BCS, reflects the body reserves available for basic metabolism, growth, lactation and reproductive activity (
13). The existence of relationship between reproductive performances and BCS in cattle has been well established (
14). However, to our knowledge no previous reports have investigated the impact of BCS, especially the impact of the low or high values, on the oxidative status in the reproductive tract.
In humans, studies on the pathophysiology of unexplained infertility have incriminated oxidative stress as an underlying factor (
15) and it has been shown that ROS may contribute to the relative inferiority of oocyte quality during in vitro assisted reproductive technologies (
16). In fact, excessive ROS production are known to overpower the body’s natural antioxidant defense system, creating an environment unsuitable for normal female physiology (
17).
In dairy cows, assessing oxidative status in reproductive tract merit investigation since the metabolites of the oxidative stress participate in several procreation processes. Analyzing the oxidative stress in situations of altered body score condition could offer real opportunities to understand underlying mechanisms affecting cow fertility.
In the present study, we determined the number of follicles according to body condition score. Furthermore, components of the primary antioxidant defense system as well as indicator for oxidative damage in the reproductive tract were also evaluated.
The results of ultrasonography showed that the number of follicles is significantly affected by body condition score (BCS). Cows with low or high BCS (<3 or>3) showed less follicles than those with BCS 3. Similar results are reported previously in rat with a significant decrease in follicles number in relation to limited nutritional exposure (
18). In human, suboptimal oocyte quality was reported in obese women (
19).
Because nutrition acts at different levels of the system that controls reproduction, it also affects the pattern of secretion of GnRH and the gonadotropins in situations of under-nutrition (
20). Diskin et al. (
21) demonstrated that dietary restriction reduced dominant follicle growth rate and adversely affected the size and ovulatory fate of the dominant follicle.
The current results revealed an obvious relationship between BCS and oxidative stress in reproductive tract. It was found that ovaries in cows with BCS 3 presented the highest levels of TAS and CAT with subsequent low concentrations of TBARS compared to cows with low or high BCS.
These findings are consistent with those of Bernabucci et al. (
22) who demonstrated that dairy cows with high BCS losses are more sensitive to oxidative stress. Importantly, it is well accepted that overfeeding or underfeeding the donor cows may influence oocyte quality and consequently the success of embryo transfer (
23). In addition, the impaired oocyte quality has been associated with excessive lipid content leading to infertility (
24). In mouse, similarly, it has been found that maternal diet status of obese female alters oocyte competence and developing embryo (
25). In murine oocytes, oxidative stress may be the initiator of ageing and has the capacity to affect mitochondrial function (
26). In human, Polak et al. (
15) found a significant depletion of antioxidants in patients with unexplained infertility than in fertile women.
In the follicular fluid, both total antioxidant status and catalase activity were significantly higher in cows with medium BCS (
3). Follicular fluid is an important microenvironment that plays a crucial role in the development and maturation of oocytes. In human, the oxidant-antioxidant state of the follicular fluid and its effects on oocyte has been largely investigated (
27). It is reported that oocyte developing within a follicular fluid of enhanced antioxidant capacity might be more fertile than ones with lower levels of antioxidant (
28). Similarly, in women undergoing in vitro fertilization (IVF), it was found that oocytes with follicular fluid containing increased total antioxidants levels resulted in higher pregnancy rates (
29). As well, it has been demonstrated that follicular fluid containing higher levels of catalase and superoxide dismutase showed better fertilization and cleavage rates in patients undergoing IVF (
30). In the other hand, Kazemiet al. (
31) reported that the duration of infertility was strongly associated with decreased antioxidant defense in women follicular environment.
In the current study a perfect similarity was observed concerning the oxidative status between the ovaries and the oviductal fluid. In both tissues total antioxidant status and catalase activity were significantly higher in cows with medium BCS.
Oviduct is an active and dynamic organ which provides an adequate environment for sperm transport and capacitation, oocyte maturation, fertilization and early embryonic development (
32). The modification of oviductal conditions may affect the embryonic developmental competence (
33). In fact, oviductal environment is the specific and responsible region for the success of fertilization process (
34), particularly; that the oviduct provides the nutrients for complete balance of antioxidants for oocyte survival (
35). On contrary, increased levels of ROS have been implicated in embryonic arrest and poor fertility outcomes (
36).
When exploring total antioxidant status and catalase activity in uterus, values were significantly higher in cows with medium BCS (
3) revealing consequently a better micro-environment compared to cows with low or high BCS. It is well established that a cow cannot conceive after calving until its reproductive tract has recovered sufficiently to support another pregnancy. However, the redox status and oxidative stress in the endometrium is still poorly defined. A recent study showed that activity of antioxidants enzymes was up regulated during pregnancy in the endometrium of a normal sheep (
37). According to the authors the increase of antioxidant enzymes between the days 16 and 21 is important to prevent a possible oxidative damage in early pregnancy. Yoon et al. (
38) showed that excessive ROS reduced the embryo development rate and increased the number of apoptotic cells in embryos cultured in vitro.
The oxidative status in plasma samples was better in cows with medium BCS than in cows with low or high BCS. Investigating the oxidative status in plasma is fundamentally important to assess a physiologically altered state, to detect adverse nutritional condition or an increased disease situation. Many substrates in plasma including oxygen, polyunsaturated fatty acids, phospholipids, cholesterol are targeted by oxidation. Several studies reported positive correlations between high BCS, plasma non-esterified fatty acid (NEFA) levels, and increased incidence of health problems in dairy cattle as a consequence of accelerated BCS drop during the periparturient period (
1,
4,
39). Interestingly, Bernabucci et al. (
22) studied the association between BCS, lipid mobilization (NEFA) and the oxidative stress imbalance; they found that cows with elevated plasma NEFA levels presented higher plasma levels of reactive oxygen metabolites and lower levels of antioxidants.
CONCLUSION
The nutritional status in cattle, reflected by BCS, is an important factor affecting reproductive function and plays a pivotal role in determining fertility. The current study demonstrated that both follicles number and oxidative stress biomarkers are significantly affected by BCS. It was shown particularly that oxidative stress is associated with low or high BCS values including cows with high (>3) and low (<3) values. These findings support that oxidative stress is probably a key factor underlying the different diseases associated with altered nutritional status.

 10.2478/macvetrev-2020-0021
10.2478/macvetrev-2020-0021