Semen cooling and cryopreservation have revolutionized the field of reproductive biotechnologies. However, challenges persist in maintaining sperm quality and viability during these processes. The unsatisfactory results and the main problems are associated with low quality, viability, morphological, structural, and DNA integrity, changes in plasma membrane, ability to interact with female tract and decrease in sperm fertilization potential. The objective of this study was to determine the effect of seminal plasma proteins with different molecular weights on the kinetic parameters of dog spermatozoa during cold storage at 4 °C. The proteins were isolated from the dogs’ seminal plasma ranging between 6-200 kDa, separated in four fractions. The ejaculates (n=15) were collected from 6 heathy dogs which were pooled. Spermatozoa were extended with Tris medium alone (control) or with addition of one of the isolated protein fractions, and were then incubated at 4 °C for 2 hours. Sperm incubated with seminal plasma proteins ranging between 10-15 kDa had significantly higher total motility (37.01±2.98%), sperm progressiveness (15.97±1.91%), curvilinear line velocity (37.46±3.75 μm/s), linearity (26.18±1.00%), and straightness (45.94±2.03%) compared to the other groups incubated with higher molecular weight proteins and the control group. The findings of this study indicated that the use of certain proteins in seminal plasma can be beneficial for reducing the detrimental effect of cooling at 4 °C and preserving the viability of dog spermatozoa. It seemed that the presence of 10-15 kDa proteins from canine seminal plasma rendered the spermatozoa less amenable to the negative influence during cooling.
The fundamental problems associated with artificial insemination using cooled dog semen involve variations in storage temperatures, variable semen quality between dogs, poor spermatozoa survival rates, disturbance of sperm motility, DNA fragmentation and etc. (
1, 2, 3, 4). The low fertility of cold shocked spermatozoa are primarily result of sperm plasma and acrosome membrane damage (
2).
Low temperatures can cause capacitation-like changes (
5) and disruption of the extracellular matrix components leading to alterations in the membrane permeability to water and ions (
5, 6). Low-temperatures induce transition of the plasma membrane from cooled crystalline to gel phase (
7). According to Martínez-Barbitta and Rivera (
2), for every 10 °C decrease, cellular metabolism is reduced by 50%, and at 5 °C metabolic activity is only 10% compared to the normal metabolism at body temperature. It is known that cooled spermatozoa from 37 °C to 4 °C undergo capacitation-like changes that affect the ability of the male gametes to achieve higher fertilization rates
in vitro (
8). Plasma membrane changes have been observed in low-temperature storage of semen, and most of them are result of seminal plasma (SP) removal (
9). Various SP components have been associated with improved sperm motility and viability. The Heparin-binding protein fraction (125 μg/mL) increased linearity, straightness and velocity of spermatozoa (
10). Caballero et al. (
6) determined that some of the seminal plasma proteins belong to the large protein family of sperm adhesins. These proteins can affect sperm motility, oviduct binding, zona binding, and even fertilization. After ejaculation, specific seminal plasma proteins are adsorbed onto the sperm plasma membrane surface, and it is hypothesized that they have an important role in maintaining membrane stability until the process of capacitation occurs (
6). However, the roles and impacts of the seminal plasma and plasma proteome regarding sperm function remain largely a matter of speculation and require further studies.
This study is focused on evaluating the effects of seminal plasma proteins with different molecular weights on the sperm kinetics during the short coldstorage of dog spermatozoa at 4 °C.
MATERIAL AND METHODSSemen collection, computer-assisted sperm analysis (CASA) and seminal plasmaSemen samples (15 ejaculates) were obtained from six dogs aged 2-6 years (
Canis lupus familiaris) by manual stimulation. All ejaculates were pooled and aliquoted in 5 samples: control, F1, F2, F3, and F4. Subsequently, the ejaculates underwent computer-assisted assessment (CASA) using a Sperm Class Analyzer (SCA) (Microptic, Spain). Total sperm motility (TM, %), progressive motility (PROG, %), curvilinear line velocity (VCL, μm/s), linearity (LIN, %), and straightness (STR, %) were measured before and after sperm cooling with different seminal plasma protein fractions. Seminal plasma was obtained by double centrifugation at 300 g at 4 °C for 5 min and at 10,000 g at 4 °C for 10 min (Labofuge 400, Germany) (
11).
Gel permeation chromatography (GPC)The GPC column was calibrated with standard (Sigma, Standard Mix 10-600 kDa) and equilibrated with phosphate-buffered NaCl (NaCl- 9 g, NaH
2PO
4-0.19 g, Nа
2HPO
4x12H
2O-1.09 g, pH 7.0). One milliliter of the seminal plasma was loaded into a semi-preparative chromatography column (SRT-10-SEC-300, Sepax) at a flow rate of 6 mL/min and 1,700 psi. Following the GPC analysis, four seminal protein fractions were obtained (faction 1 - F1, fraction 2 - F2, fraction 3 - F3, and fraction 4 - F4). Each fraction contained proteins with different molecular weights.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE)
The GPC protein fractions were characterized by 15% SDS-PAGE (BioRad Mini Protean Tetra System), and the Plus One Silver Staining Kit (GE Healthcare) was used for visualization. The molecular weight of the four protein fractions varied between 6-7 kDa and over 200 kDa, as follows: fraction 1 included bands with proteins with high molecular weights (55-60 kDa, 74-78 kDa, and 180-200 kDa), fraction 2 contained bands with proteins with molecular weights 45-60 kDa, bands with molecular weights 17-35 kDa, fraction 3 contained band with low molecular weight (14-15 kDa), and band with proteins of 10-11 kDa, and fraction 4 contained band with proteins of 6-9 kDa.
Incubation of samples with GPC fractions at 4 °CAfter an initial CASA assessment, the semen samples were mixed and centrifuged at 300g for 5 min to exclude the SP (Labofuge 400, Germany). Following centrifugation, the semen was resuspended in a medium (Tris - 3.025 g, Citric acid - 1.7 g, Fructose - 1.25 g), at a ratio of 1:2. The re-suspended semen samples were aliquoted into volumes of 800 μL, and 200 μL of each seminal protein fraction was added into the samples, resulting in a final volume of 1,000 μL. The sample with seminal plasma (not extracted from the sample) was used as a control and it was re-suspended only with the medium. All samples and control were cooled at 4 °C for 2 hours. The effect of the various seminal protein fractions (ranging from 6 to 200 kDa) on sperm kinetics were measured and compared at the 1
st and 2
nd hours of cooling at 4 °C.
Statistical analysisThe results are presented as mean values ± standard deviation (SD). The data were analyzed using T-test in Excel. Significance level was set at р<0.05.
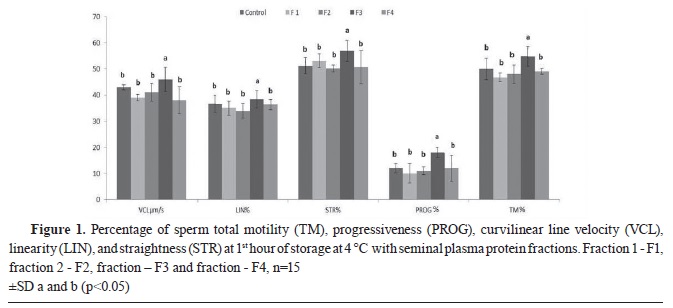
RESULTSThe fresh ejaculates had TM-72.05±6.69, PROG-26.09±2.69, VCL-71.01±4.19, STR-64.18±1.00, and LIN-49.08±4.54. The observed CASA parameters at the 1
st and 2
nd hours of cooling with addition of different seminal plasma protein fractions are presented in
Fig. 1 and
Fig. 2.
The TM of spermatozoa decreased (p<0.05) in all observed samples and in the control after the 1
st h of storage at 4 °C (
Fig. 1). No significant differences in sperm TM were observed among the control (49.82±1.54), F2 (47.11±2.05), and F4 (48.54 ±2.46) The lowest decline of observed sperm parameters was observed in F3 samples compared to the other groups. The sperm PROG exhibited significantly higher values in F3 compared to the other groups. The lowest PROG and TM values were observed in F1 samples (PROG-10.08±1.03 and TM-43.77±3.35) compared to F3 (PROG-19.96±2.26 and TM-54.88±1.03). STR for F3 samples (56.92±2.18) was significantly higher compared to the F2 (50.19±1.11), F4 (50.66±6.88), and the control (51.24±5.80) after the 1
st hour of storage. VCL (46.94±2.23) and LIN (38.99±1.52) for F3 were significantly higher compared with the control and the other groups at 1
st h storage.
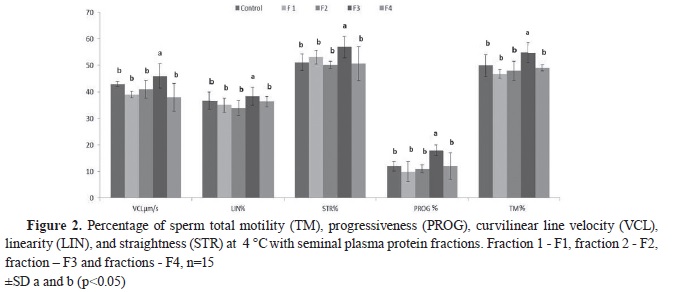
However, the most significant changes in the observed parameters were established during the 2
nd hour (р<0.05) of storage at 4 °C in all samples and the control (
Fig. 2). The experiment demonstrated that F3 samples maintained higher values of PROG (15.97±1.91) and TM (37.01±2.98) compared to other samples (F1 PROG (8.09±1.84), TM (43.17±1.39); F2 PROG (9.11±1.66), TM (44.36±1.15); F4 PROG (11.46±1.12), TM (46.75±1.41); and the control PROG (11.37±1.18), TM (48.16±2.05). A significantly lower VCL was registered in F1 (29.03±1.67), F2 (31.12±2.31), F4 (27.44±1.87), and the control (32.65±1.98) compared to F3 (37.46±3.75).
After the 2
nd hour of storage at 4 °C, F3 samples exhibited higher percentage of LIN (27.18±1.00), and STR (46.94±2.03) compared to the other samples and the control (for F1-LIN–19.63±2.00 and STR–40.66±1.15; for F2-LIN-21.61 ±1.00 and STR-41.02±2.92, for F4-LIN-22.97±3.05 and STR-39.94±.4.44, and for control-LIN-19.99±2.45 and STR-39.83±1.87).

 DISCUSSION
DISCUSSIONThe low-temperature storage of semen often alters the functionality of spermatozoa by destabilizing the plasma membranes of the cells subsequently leading to impairment of their main functions (
12). The effect of seminal plasma proteome on sperm functionality remains largely a matter of discussion (
13, 14, 15, 16). Some authors reported that the addition of canine zinc-binding proteins (ZnBP) isolated from the seminal plasma has a protective effect on spermatozoa motility and membrane integrity during 5 °C storage (
12). Mogielnicka-Brzozowska et al. (
16) demonstrated that ZnBP in canine seminal plasma with molecular weight of 10.7 kDa and 14.2 kDa might be implicated in key events related to reproductive processes in dogs. Furthermore, in other species, certain ZnBP in seminal plasma has demonstrated positive effects on sperm viability, motility, mitochondrial activity, and intact acrosomes after storage at 5 °C (
6, 17). During conditions of cold shock, spermatozoa absorb eight times more zinc (Zn) than at physiological temperatures (
18). It is known that excessively high Zn content exerts an inhibitory effect on sperm motility (
19). Moreover, the mechanism of ZnBP action on canine spermatozoa motility may involve their ability to chelate free or loosely-bound zinc ions, which prevents their excessive absorption by spermatozoa under cold shock conditions. Such a property was observed in human ZnBP (
20). Some authors noted that the addition of prostatic fluid may have a positive effect on the fertilization capacity of extended and cooled canine spermatozoa (
2, 21). This study aimed to determine GPC separated seminal plasma proteins that could have a positive effect on canine spermatozoa cooled at 4 °C. In our experiments, we observed canine seminal plasma proteins with molecular weight between 10-15 kDa, which may have a potential effect on preserving the sperm kinetic parameters. The acquired data shows that after the incubation of spermatozoa with proteins with different weights at 4 °C, fraction 3 containing low molecular weight proteins has significantly favorable effects on sperm TM, PROG, LIN, STR, and VCL, compared to high molecular weight seminal plasma proteins. However, the effect of adding SP and seminal plasma proteins on sperm functions has been widely studied and remains a subject of research (
22, 23). Low molecular weight seminal plasma proteins (3 kDa) prevent deleterious effects when added before decreasing the temperature to 5 °C, increasing sperm viability from 26% to 34% while preserving membrane integrity, thus indicating a protective effect against cold shock (
14). According to Bubenickova et al. (
10), the role of SP as is still unclear because it can either stimulate or inhibit the functions of male gametes. Seminal plasma appears to have a protective role during
in vitro manipulation of spermatozoa in some species, as processes such as cooling not only affect motility and viability, but also the ability of spermatozoa to penetrate the cervix during the insemination (
22, 24). The addition of SP during cooling has been found to exhibit contradictory results, depending on the mammalian species. The role of SP varies significantly in terms of its effect on spermatozoa, particularly within the context of
in vitro supplementation during semen storage. However, the reasons for these differences are yet to be determined along with the complete role of seminal plasma in the biological function of the spermatozoa (
25). Promising results have been reported by Pan et al. (
23) which used an extender with SP concentration of 25% leading to effective preservation of the canine sperm motility and plasma membrane integrity in samples chilled at 4 °C. The inclusion of SP proteins with a molecular weight of 14 kDa and 16 kDa and isoelectric points between 5.0-5.5 in the preservation medium increased bull sperm viability (
26). The use of seminal plasma proteins in the range of 12–20 kDa leads to improvement of biotechnological preservation of spermatozoa (
27). An isolated fraction from ram SP is capable of reversing the effects of cold shock. The inclusion of a low molecular weight SP protein (approximately 20 kDa) to the medium is encouraging and has the capacity to reverse capacitation-like changes during storage at low-temperature, assuming that the adsorption of this protein to cold-shocked spermatozoa restores membrane permeability (
28). According to other authors, integrity and motility can only be regenerated when SP is ultra-filtered, and a seminal protein fraction with a molecular weight below 50 kDa is added to the medium (
29). Similarly, there was a linear correlation between motility in lowtemperature storage stallion spermatozoa and the relative concentration of two protein bands within the range of 19.6 and 15.3 kDa (
30). However, whether the protein fractions affect the membrane structures and also sperm viability or they could alter the sperm kinetics through increased energy demand, remains unclear and further research in this area is needed. Low-temperature storage is an important method in reproductive biotechnologies across species. Identifying the proteins contained in SP should establish better protocols for semen storage for reproductive assistance. Our experiments provide valuable information on the effect of seminal plasma proteins with different molecular weights in maintaining the kinetic characteristics of canine spermatozoa subjected to cooling at 4 °C. The presented data underline the need for further research into the role of seminal plasma proteome and its effects on cooled dog spermatozoa. Extensive further characterization of the canine seminal plasma proteome is essential to fully comprehend the purpose, function, and potential of seminal plasma.
CONCLUSIONThe use of seminal plasma proteins within the range of 10-15 kDa in cooled dog sperm, incubated for 2 hours at 4 °C, preserves the percentages of sperm PROG, TM, LIN, STR, and VCL parameters when added to the medium. Our results emphasize the significant role of seminal plasma proteins and their inclusion in preservation techniques for maintaining sperm viability.
CONFLICT OF INTERESTThe authors declare that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
ACKNOWLEDGMENTSThis research was supported by the Institute of Biology and Immunology of Reproduction “Acad. Kiril Bratanov”- Bulgarian Academy of Science.
AUTHORS’ CONTRIBUTIONSTsSTs performed semen collection, made a primary CASA-assessment of the canine ejaculates and conducted Gel permeation chromatography for the separation of fractions. DBD performed the SDS- PAGE, the incubation of the chromatographically separated fractions with sperm samples and the statistical analysis.

 10.2478/macvetrev-2023-0027
10.2478/macvetrev-2023-0027