The bone morphogenetic protein 15 (BMP15) and growth differentiation factor-9 (GDF9) belong to the transforming growth factor beta (TGFβ) family. They are potent regulators of ovarian functions (
2,3). In the available literature, there are no sufficient data about the expression of bone morphogenetic proteins and their role in hens’ ovary. The studies primarily focus on BMP4 and BMP7 (
4). Oocyte competence in egg laying species is very important because the proper yolk accumulation in egg is closely coordinated by steroidogenesis and follicular maturation. Preovulatory follicles are classified according to their size. The ovulation occurs with the largest follicle which precedes the second and so forth. There are not enough data on the early stages of laying hens folliculogenesis (
5). Hypothetically, BMP15 may be specific in regulating the follicles and reproductive processes in poultry. The aim of the present work was to determine BMP15 mRNA expression and its receptors (BMPR1B and BMPR2) in laying hens’ ovary around the time of follicle selection.
MATERIAL AND METHODSAnimalsThe current experiment was conducted with forty laying hens from Lohman Klassik Brown breed (40 weeks old). Layers were randomly divided into four replications, 10 poultry each. The hens from each replication were raised in separate boxes on a deep litter pen. The poultry received
ad libitum water through nipple watering troughs and 130 g/day/hen compound feed for laying hens. The diet contained maize, sunflower meal, soybean meal, sunflower oil, salt, limestone, monocalcium phosphate, L lysine, mineral premix, vitamin premix, and Sinergin®. The compound feed had the following chemical composition: 11.05 MJ metabolizable energy, 16.7% crude protein, 4.3% crude fats, 3.5% crude fiber, 12.14% crude ash, 3.70% Ca, 0.47% available P. The trial duration was 50 days.
Collection of experimental materialsThe experimental protocol used in this study was approved by the Ethical committee of the Institute of Animal Science - Kostinbrod, Bulgaria and its protocol has been approved by the National Ethics commission for animals within the delivered permission of use of the animals in the experiments (N85/04.10.2018, expiry date 04.10.2023). At the end of the trial fifteen layers were randomly selected and humanely killed in accordance with Directive 2010/63/EC of the European Parliament (
6).
Hens’ ovaries were then dissected, rinsed with PBS and transported in storage medium (pH=7.1) at room temperature to the lab of Institute of biology and immunology of reproduction “Acad. K. Bratanov”, BAS. Follicles larger than 2 mm (size 3-5 mm) were mechanically removed from the ovaries. Experimental materials were obtained from follicles with size 6-8 mm and bigger than 9 mm. The oocytes were cleared of granulosa cells according to the methodology of Gilbert et al. (
7). Samples of ooplasm and granulose cells were collected individually (n=15 of each).
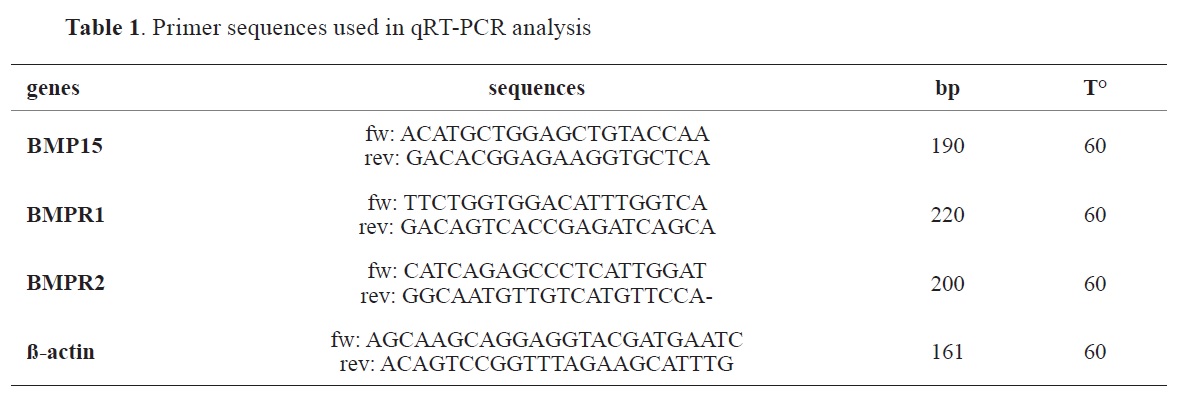
Quantitative Real-Time PCRRNA was extracted from the described samples, using RNA easy Mini Kit with a column (Qiagen Inc., USA). Reverse transcription was done with 1 μg of RNA template with a SuperScript II RNase H Reverse Transcriptase kit (Invitrogen, USA). Each sample was processed in triplicate using a cycler real-time PCR instrument (Agilent Stratagene Mx3005P) and involved a total volume of 20 μL, including 500 nM primers with the indicated sequences (
Table 1).
Real-time PCR reactions were processed at 95 °C for 5 min, 40 cycles of 95 °C for 15 s, 60 °C for 1 min, 95 °C for 15 s, 60 for 1 min and 95 °C for 15 s. The ß-actin gene was used as a reference. Mean and standard deviation (SD) of the relative gene expression were evaluated using the 2(-delta delta C(t) method (
8).
Western blotProtein lysates were mixed between two individual hens (n=10) and after the samples were supplemented with 2 X Buffer and loaded (15 μl each) on 10% SDS polyacrylamide gel under reducing conditions. Protein fractions were visualized by Coomassie Brilliant blue (Sigma Co., USA) and their molecular weight was determined according to standard molecular markers. Proteins were transferred to a nitrocellulose membrane (Millipore, USA) for 1 hour. Nonspecific binding sites were blocked with 5% nonfat dry milk in TBS (pH=7.6) for 1 hour at room temperature. The blots were incubated overnight at 4 °C with a primary antibody - rabbit against chicken BMP15 (Santa Cruz Biotechnology, CA) at a concentration of 1:750 in blocking solution. After extensive washing with TBS, goat anti-rabbit HRP-conjugated second antibody (Santa Cruz Biotechnology, CA) was added in a concentration of 1:5000 for 1 h at room temperature. Immuno-detection of proteins was revealed using alkaline-phosphatase buffer (pH=9.5) in which nitro blue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolyl-phosphate (BCIP) were added. The reaction was stopping in dH
2O.

*Note: A correction of a sentence is being made in the Materials and Method part. Please see https://macvetrev.mk/LoadAbstract?DOI=_
StatisticsStatistical analysis was performed using the software program StatSoft, version 10 (StatSoft Inc., Tusla, USA). The differences between different samples were analyzed using the Student’s t-test. Values are presented as means ± SEM and were assumed to be statistically significant at p<0.05.
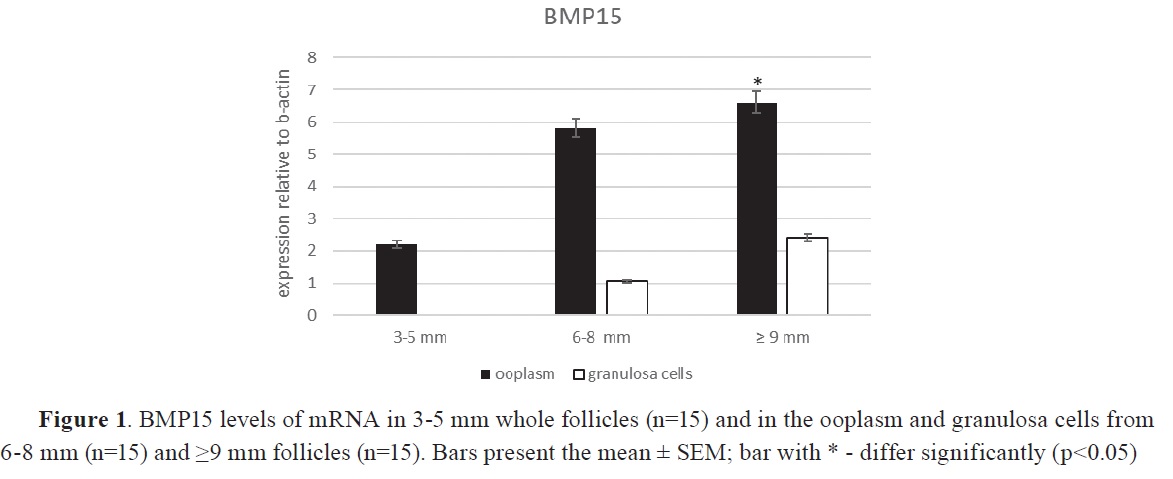
RESULTSSignificantly higher levels of BMP15 mRNA (p<0.05) were detected in ooplasm compared to granulosa cells, regardless of follicle size by real time PCR analysis. The expression was significantly higher (p<0.05) in oocytes from mature follicles of 6-8 mm (3.86-fold) and ≤9 mm (4.30-fold) diameter compared to smaller follicles, with equal numbers between the three groups (n=15), (
Fig. 1).



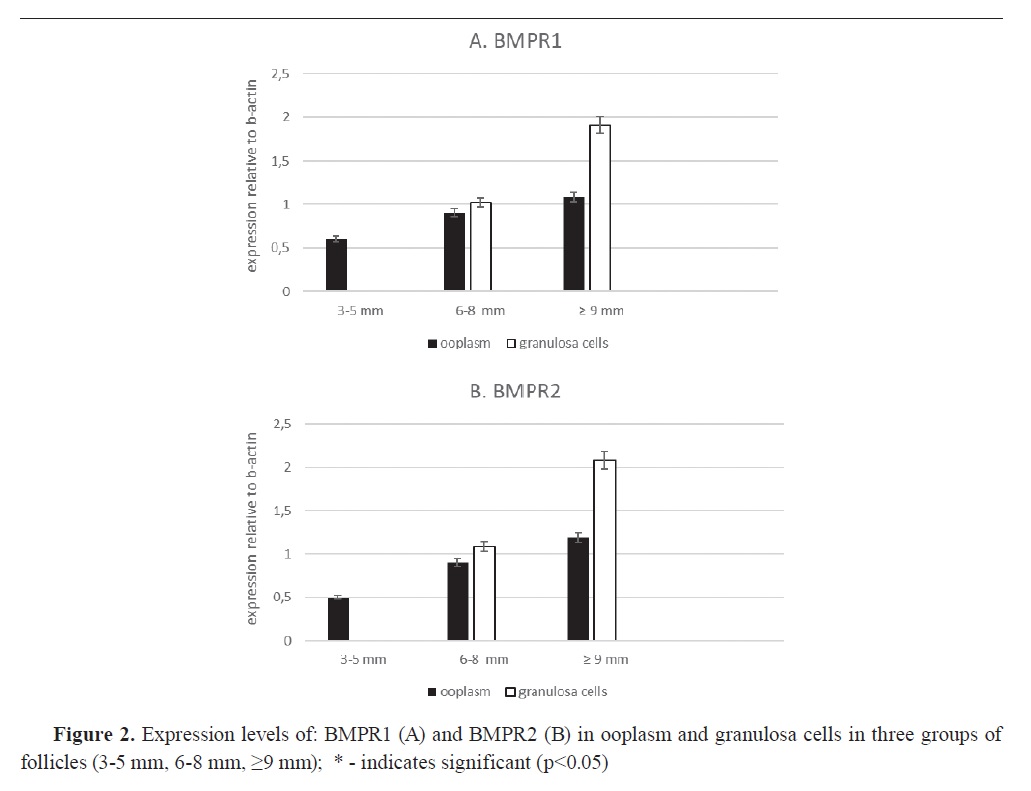
In the period of follicle development, PCR analysis showed that mRNA for both BMPR1 (
Fig. 2A) and BMPR2 (
Fig. 2B) was significantly higher (p<0.05) by 1.15 and 1.71 times compared to mRNA expression in 3-5 mm follicles. Expression signals between both receptors were approximately the same in the prospective follicular phases. The largest follicles (≥9 mm) had two-fold higher receptor expression levels in the granulosa cells compared to the secondary follicles (6-8 mm) (p<0.05), which suggests localization of these receptors mainly in granulosa cells.
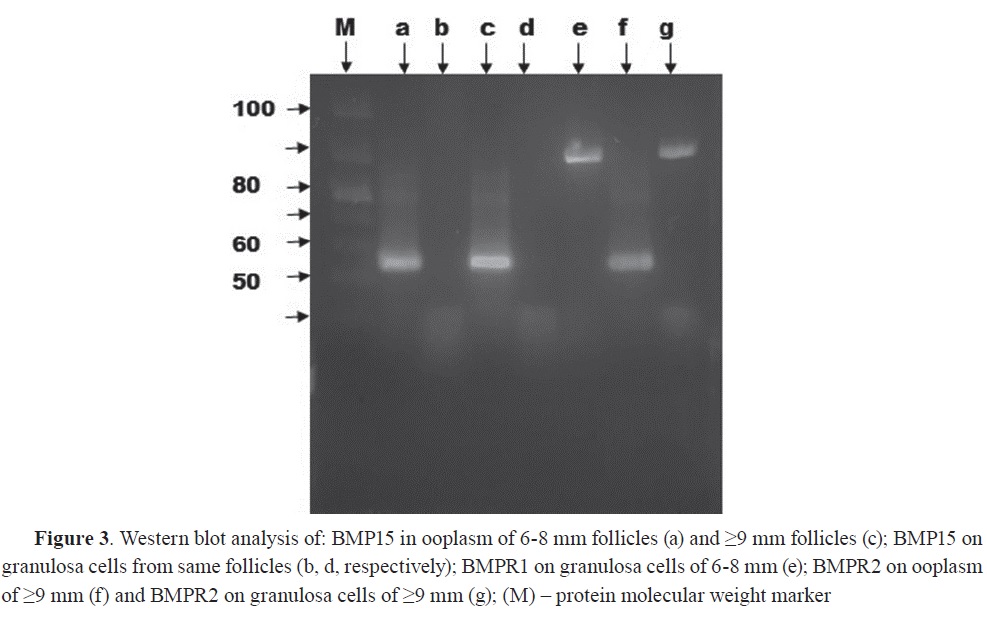
Polyclonal anti-BMP15 antibody recognized protein band corresponding to approximately 55 kDa molecular weight in both samples of ooplasms of secondary (6-8 mm) and large (≤9 mm) follicles (
Fig. 3, a and c). This band was not found in the granulosa cells which matches the result obtained by PCR (Fig. 3, b and d). BMPR1 and BMPR2 were detected with Western blot at around 90 kDa in granulosa cells of 6-8 mm follicles (
Fig. 3, e) and ≥9 mm follicles (
Fig. 3, g) with small detectable concentration in the ooplasm (
Fig. 3, f).
DISCUSSIONThis is one of the few articles documenting BMP15 and its receptors expression, function and regulation during the folliculogenesis in hens. In this study, 3 mm follicles, ooplasm, and granulose cells were used for detection of BMP15 mRNA expression. The current results comply with the study of Elis et al. (
9) who reported that BMP15 mRNA expression was predominantly in the oocyte whereas its receptor was in the granulosa cells. Their signal was positively correlated with the follicular development stage. Compared to mammalian species, the main source of BMP15 in the ovary was the oocyte (
10).
A single band approximating 50 kDa was present in the ooplasm of 6-8 mm follicles, but the protein signal was undetectable in the granulosa cells and in small follicles. In contrast, Western blot analysis proved the presence of BMP15 protein in ovine follicular fluid (
11). The results from the current experiment were similar to findings in seabass, where signal of BMP15 mRNA was localized during early stages of follicle development, but protein expression wasn’t detected until the beginning of secondary follicular growth phase (
12).
Increased expression of BMP15 mRNA and protein was also detected in the larger rat follicles by
in situ hybridization and immunohistochemistry (
13). This might indicate that BMP15 expression in granulosa cells is species dependent.
The predominantly higher expression of mRNA for BMP15 receptors - BMPR1 and BMPR2 is in the granulosa cells of the larger follicles. Higher expression is in positive correlation with the follicle development in hens and could be used as an indicator. BMP15 receptors have support function in the signaling between the oocyte and the granulosa cells (
14).
The small follicles (≤3 mm) in the hen ovary grow slowly until they are selected into the preovulatory period. The oocyte size increases drastically after selection due to yolk accumulation. If not selected, they undergo atresia. Although, mechanisms of selection of dominant follicles remain unknown, many authors point that FSH and AMH are main factors for selection (
15, 16). Members of BMPs group have an effect on the signaling function of FSHR and AMH in hens, thus contributing to the normal follicular development (
17, 18).
CONCLUSIONThe results obtained in this study demonstrate the possible role of BMP15 on follicular development in hens’ovaries. BMP15 expression is important for the growth regulation and signaling in the follicular cells at the preovulatory stage, granulosa cell differentiation, increased BMPR1 and BMPR2 expression, and yolk formation, preparing the oocyte for ovulation.
CONFLICT OF INTERESTThe authors declare that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
ACKNOWLEDGMENTSThis research was supported by the Ministry of Education and Science of Republic of Bulgaria, Project DM 16/4.
AUTHORS’ CONTRIBUTIONSSJG has designed the biological experiment. DVA conducted the laboratory work and interpretation of the results. The manuscript was drafted and reviewed by DVA and SJG.

 /10.2478/macvetrev-2023-0026
/10.2478/macvetrev-2023-0026