Mac Vet Rev 2015; 38 (2): 183 - 188
 10.14432/j.macvetrev.2015.06.048
10.14432/j.macvetrev.2015.06.048
Received: 12 May 2015
Received in revised form: 15 June 2015
Accepted: 19 June 2015
Available Online First: 26 June 2015
Published on: 15 October 2015
Keywords: sheep, season, metabolic profile
Ruminants have different carbohydrate metabolism compared to non-ruminant animals. Namely, dietary glucose in sheep and other ruminants is converted into short-chain volatile fatty acids in the rumen, so the main source of blood glucose is gluconeogenesis in the liver (1), with gluconeogenesis evident even in the fetal sheep liver (2). This also causes blood glucose concentrations to be lower than in non-ruminant animals, and predispose the animal to a negative energy balance (NEB), when energy demands arise. In pregnant ewes, the increased fetal demand is not combined with increaseed endogenous glucose production, so maternal tissues manifest a relative energy deficit, compared with non-pregnant ewes (3). This energy deficit causes mobilization of fatty acids from the adipose tissue which are utilized in the liver, leading to increased level of ketone bodies, especially beta-hydroxibutirate (BHBA) (4). This condition could progress into a very severe metabolic disorder - pregnancy toxemia (5), a disease manifested with neural signs due to the fact that glucose is the only energy source for the brain (6). There are various factors which have an impact on energy metabolism and contribute to the development of a negative energy balance (NEB). Those are food intake, starvation (7,8), hormonal status (9,10,11,12,13,14), age and gender (15), twin-bearing (16) and cold stress (17). Pregnancy in sheep commonly occurs during the winter season when animals might be exsposed to cold stress. Exposure to low enviromental temperatures decreases insulin secretion (17). In the condition of a disturbed energy balance, some biochemical parameters in the serum, which are indicators of metabolic disorder and hepatic insufficiency could be changed. These parameters could also be changed due to exposure of animals to extreme enviromental temperatures, as was found in cattle (18). Therefore, the aim of this study was to determine possible seasonal variations of biochemical parameters in Chios sheep.
Multiparous Chios ewes from one flock in Pelagonia, a region in R. of Macedonia, were chosen for the study. One group of ewes (n=8) were chosen during the early winter season (December), when cows were exposed to cold stress, while the other group (n=8) was chosen during the spring season(May). Ewes examined during the winter season were pregnant and manifested clinical signs of pregnancy toxemia (inappetence, ataxia, lethargy, weight loss) and ewes investigated during the spring season were non-pregnant and clinically healthy. During the winter period, the flock was exposed to enviromental low temperature and consumed frozen low quality roughage hay, that caused many lesions in the oral mucosa. According to the data from the flock farm, ewes were not treated for endoparasites. Blood samples were taken from each animal included in study, by venepunction from v. jugularis. Biochemical parameters that were measured in the blood samples were glucose, BHBA, total protein, albumin, urea, creatinine, triglycerides (TG), cholesterol, AST and ALP. Glucose, total protein, albumin, triglyceride, cholesterol were determined by standard “end point” methods according to the manufacturer’s instructions Human, Germany. AST, ALP (Human, Germany) and BHBA (Randox, United Kingdom) were measured by kinetic methods according to the manufacturer’s instructions, on semiautomatic photometer STAT Fax 3300 (Awareness Technology, Inc, USA).
The obtained results for concentrations of glucose, BHBA, total protein, albumin, urea, creatinine, triglycerides and cholesterol, and for activity of AST and ALP in examined ewes are shown in Table 1.
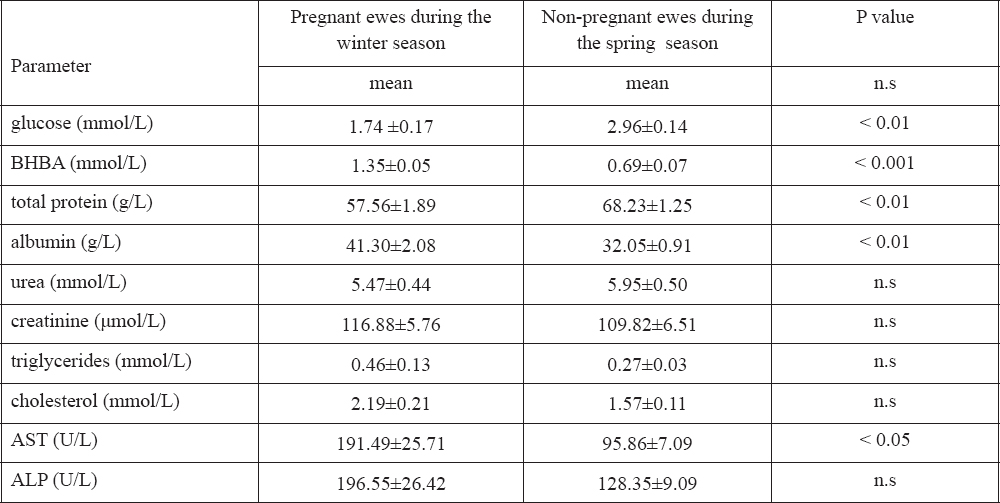
Table 1. Concentrations (X±SE) of some biochemical parameters in ewes during different seasons. P values indicate the seasonal effect
According to the results, the group of pregnat ewes during the winter session had significantly lower level of glucose and total protein (1.74 ± 0.17 mmol/L, and 57.56 ± 1.89 g/L, respectivey) than non-pregnant ewes during spring session (2.96 ± 0.14 mmol/L, and 68.23 ± 1.25 g/L, respectively) (p < 0.01), and significantly higher level of albumin (41.30 ± 2.08 g/L vs. 32.05 ± 0.91 g/L, respectively) (p < 0.01). BHBA levels were significantly higher in pregnant ewes during the winter season (1.35 ± 0.05 mmol/L) than in non-pregnant ewes during the spring season (0.69 ± 0.07 mmol/L) (p < 0.001), and also this was found in levels of AST (191.49 ± 25.71 U/L in pregnant ewes vs. 95.86 ± 7.09 U/L in non-pregnant ewes) (p < 0.05). There were no significant differencies in values of urea, creatinine, triglycerides, cholesterol and ALP between two groups.
Pregnant ewes have high energy demands due to intensive fetal growth during the last few weeks of pregnancy (14). Pregnant sheep exposed toextremly low environmental temperatures have to increase generation of heat for maintaining body temperature. This could be obtained by intake of a high-energy diet (19). Due to increased energy demands of pregnant ewes, adaptation to environmental conditions could be compromised, especially if cold exposure is accompanied with fasting. Common metabolic disorder in late pregnancy in sheep, especially in sheep with two or more lambs, known as gravidity toxemiais usually caused by metabolic imbalance and environmental stress (5).
Many investigations with sheep exposed to cold (17, 19, 20, 21, 22) have shown that extrimely low enviromental temperature may causes hyperglycemia, due to decreased secretion of insulin from the pancreas (21). This is opposite toour results, which showed significant hypoglycemia in sheep with pregnancy toxemia during the winter season, when animals were exposed to extrimely low enviromental temperature and decreased feed intake. Contrary to this, during the spring period animals showed normoglycemia, which is in accordance with the results obtained on sheep exposed to normal enviromental conditions (17). There was significant difference (p<0.001) in the glucose level between ewes exposed to cold stress (1.74 ± 0.79 mmol/L) and ewes not exposed to cold stress (2.96 ± 0.44 mmol/L). This finding obviously differs from results of others, probably because the latter included fattering breeds of sheep Suffolk (17, 19, 20), Suffolk-Hampshire-Rambouilet crossbred (21), Corridale-Suffolk crossbred (22), respectively. This suggests possible significant differences in energy metabolism in dairy and fattering breeds of sheep. In pregnant ewes gluconeogenesis does not increase with increased fetal demand, which may causes energy deficit of maternal tissues. This is not a case in non-pregnantewes (3). Low concentration of serum glucose inpregnant ewes is probably the consequence of inappropriate gluconeogenesis in the liver from glucogenoplastic precursors, like propionate that originate from rumen fermentation. Food deprivation, which was evident in pregnant ewes during the winter season, causes lack of propionate precursors for glucose production. Biochemical pathways are further complicated with inappropriate beta-oxidation of acetate and butyrate leading to increased production of acetil-Co-A by hepatocytes. This product is a precursor for aceto-acetyl-Co-A. Aceto-acetyl-Co-A is a primordial molecule for ketone bodies with high effect on acido-base status. This molecule is converted into a very unstable molecule acetoacetate. Reduction of acetoacetate produces BHBA, while irreversible decarboxilation of acetoacetate produces aceton molecules. Excessive concentration of serum BHBAcannot be utilized by extrahepatic tissue, and therefore the serum level of BHBA remainshigh. Significantly marked hyperketonemia was noticed in pregnant (1.35 ± 0.27 mmol/L), but not in non-pregnant sheep (0.69 ± 0.21 mmol/L), though it was not as high as found by other authors (24). The difference in BHBA concentrations between pregnant and non-pregnant sheep was significant (p<0.001), due to decreased alimentary energy resources in pregnant sheep. BHBA is an indirect indicator of negative energy balance, and season variation during the late spring period showed lower serum concentration as a result of better energy supply.
Insufficient energy derived by carbohydrate metabolism causes lipolysis, but energy derived from free fatty acids cannot be adequatelly used in Krebs cycle, because of a lack of oxalacetate, derived from glycogenic precursors. The hepatic capacity for metabolizing of non-esterified fatty acids (NEFA) released from adipose tissue is overloaded. Physiologicaly, NEFA can be reesterified in triglycerides which are included in the formation of VLDL for secretion in blood. Sheep have a very low capacity for production of VLDL (24), especially breeding and non-breeding sheep compared with sheep in lactation (13), and thereby not capable to produce enough VLDL in condition of compromised lipid metabolism in hepatocites. In that case, TG can not be removed from hepatocytes, remaining in the hepatocytes causing hepatic insufficency. In our study, this was evident by higher serum activity of AST (191.49 ± 117.84 U/L) in pregnant ewes which suffered from severe liver impairment and had decreased liver capacity to maintain gluconeogenesis, compared to non-pregnant ewes (128.35 ± 27.27 U/L) (p<0.05). AST is a liver enzyme that enhances the process of transamination, with oxalacetate, a main metabolite in Krebs cycle, as a final product. As a cytosol enzyme with many isoformes it can easily passthe hepatocyte membrane, so increased serum activity of AST indicates an over loaded hepatocyte activity. Thus, in our study the liver was affected by metabolic distress during the critical period and further complicated by lack of alimentary glycogenic precursors. Homeostatic regulatory mechanisms are disrupted, because of neuro-endocrinological requirements in the period of late gestation and lambing. This critical period in the sheep is further complicated with low environmental temperature and high metabolic energy demand. Variations in environmental conditions during the cold season reflects on the energy metabolism, presented through biochemical parameters. So environmental stress, inappropriate feeding management and inappropriate dehelmintisation decompensate liver function and capacity to survive the critical period. Stress hormones, such as high values of serum cortisol level, further enhance the catabolic processes in the extra mammary tissue and skeletal muscles, affecting the body condition score. It is known that pregnancy toxemia is accompanied with increased cortisol level (24).
Particularly, serum protein fractions pattern could give information about dehydration, plasma volume expansion and hepatic function. Particularly, serumprote infractions pattern could give information about dehydration, plasma volume expansion and hepatic function.
Another indicator of liver dysfunction are serum proteins levels, especially albumin, which decrease during hepatic insufficiency. In our study, the albumin level was significantly higher during the winter period in pregnant sheep (41.30 ± 9.53 g/L) compared with non-pregnant sheep (32.05 ± 2.73 g/L) (p<0.001), but in fact this was probably pseudo hyperalbuminemia, as the result of dehydratation, which is related with changes in particular serum protein fractions (25). Dehydraton of sheep appears as a result of the osmotic activity of ketone bodies, because they are very strong acids causing keto-acidotic condition. Lack of water consumption also is one of the main reason for dehydration. Serum concentration of total proteins (68.23 ± 3.76 g/L) showed significantly higher values in non-pregnant sheep during the late spring period, because of regular alimentary protein precursors supply, compared to underfed pregant sheep (57.65 ± 5.44 g/L). Status of proteinemia clearly revealed appropriate synthesis and utilization when amino acids precursors are supplied from alimentary recourses. Statistically significant lower concentration of total proteins (p<0.001) in pregnant ewes during the winter period is a result of proteolysis, caused by energy deficit.
There was no significant difference between serum concentrations of urea, creatinine, cholesterol and ALP in pregnant sheep during the winter period (5.47 ± 2.04 mmol/L; 116.88 ± 26.41 µmol/l; 2.19 ± 1.00 mmol/L, 196.55 ± 121.10 U/L, respectively), and non-pregnant sheep during the spring period (5.95 ± 1.51 mmol/L; 109.82 ± 19.54 µmol/l; 1.57 ± 0.35 mmol/L; 128.35 ± 27.27 U/L respectively), and these values were all within physiological ranges (26). Also, serum concentrations of triglycerides (0.46 ± 0.60 mmol/L) in pregnant sheep showed mild decrease of values, probably due to reesterification of triglycerides in hepatocytes, but it was not statistically significant compared with non-pregnant sheep (0.27 ± 0.10 mmol/L).
Metabolic pathways in pregnant ewes were not able to overcome the negative energy balance during the winter period. On the contrary, the spring period favorably affected the energy status of Chios sheep. During the winter period, the Chios sheep were in an inadequate energy status during late pregnancy and lambing when poor feeding management occurred. Glucose and BHBA concentrations could be reliable indicators of negative energy balance and gravidity toxemia in Chios sheep.
Copyright
© 2015 Dzadzovski I. This is an Open Access article distributed under the terms of the Creative Commons Attribution-Non Commercial License (http://creativecommons.org), which permits unrestricted non-commercial use, distribution, and reproduction in any medium provided the original work is properly cited.
Conflict of Interest Statement
The authors declared that they have no potential conflict of interest with respect to the authorship and/or publication of this article.
Citation Information
Macedonian Veterinary Review. Volume 38, Issue 2, Pages 183-188, p-ISSN 1409-7621, e-ISSN 1857-7415, DOI: 10.14432/j.macvetrev.2015.06.048, 2015